|
Арктика и Антарктика
Правильная ссылка на статью:
Васильчук А.К., Васильчук Ю.К. Палинологическая характеристика едомы Зеленого Мыса и норок сусликов // Арктика и Антарктика. 2021. № 1. С. 26-46. DOI: 10.7256/2453-8922.2021.1.32985 URL: https://nbpublish.com/library_read_article.php?id=32985
Палинологическая характеристика едомы Зеленого Мыса и норок сусликов
Васильчук Алла Константиновна
доктор географических наук
ведущий научный сотрудник, Московский государственный университет им. М.В. Ломоносова (МГУ)
119991, Россия, г. Москва, ул. Ленинские Горы, ГСП-1, 1, географический факультет, НИЛ геоэкологии Севера
Vasil'chuk Alla Constantinovna
Doctor of Geography
Leading Research Fellow, Laboratory of Geoecology of the Northern Territories, Faculty of Geography, Lomonosov Moscow State University
119991, Russia, g. Moscow, Leninskie Gory, GSP-1, 1,, geograficheskii fakul'tet, NIL geoekologii Severa

|
alla-vasilch@yandex.ru
|
|
 |
Другие публикации этого автора
|
|
|
Васильчук Юрий Кириллович
доктор геолого-минералогических наук
профессор, кафедра геохимии ландшафтов и географии почв, Московский государственный университет имени М.В. Ломоносова, географический факультет
119991, Россия, г. Москва, ул. Ленинские Горы, 1, оф. 2009
Vasil'chuk Yurij Kirillovich
Doctor of Geology and Mineralogy
Professor, the department of Geochemistry of Landscapes and Geography of Soils, M. V. Lomonosov Moscow State University
119991, Russia, g. Moscow, ul. Leninskie Gory, 1, of. 2009
|
|
vasilch_geo@mail.ru
|
|
 |
|
DOI: 10.7256/2453-8922.2021.1.32985
Дата направления статьи в редакцию:
25-05-2020
Дата публикации:
13-04-2021
Аннотация:
Палинологическая характеристика отложений разреза Зеленый Мыс фиксирует примерно среднепериодные циклические изменения растительного покрова, как в локальном, так и в региональном масштабе. Палинологическая характеристика разреза едомных отложений Зеленый Мыс дает основание для выделения на диаграмме трех локальных палинозон: Varia +Artemisia, Varia + Selaginella sibirica и Pinus + Betula sect. Nanae. Практически все палиноспектры отражают открытые ландшафты мозаичных гипоарктических тундр. Палинозоны, выделенные по результатам палинологического повторно-жильных льдов, соответствуют палинозонам, вмещающих льды отложений: Varia + Selaginella sibirica датируется 41.9 – 33.8 тыс. кал. лет, Pinus + Betula sect. Nanae (P+B) приблизительно датируется 33.5–30 тыс. кал. лет назад. В пределах выделенных палинозон преимущественно сохраняется характер циркуляции атмосферы в весенне-летний период, циклы отражают изменения базиса эрозии и колебания температур вегетационного периода, а также его длительность. Состав региональной составляющей палиноспектров в норках и синхронных им горизонтах практически не отличается. Исключением является заметное количество пыльцы лиственницы (10%) в ископаемой норке в едоме Зеленого Мыса, это показало, что в норках создаются благоприятные условия для сохранения пыльцы лиственницы. Семена, обнаруживаемые в норках, отражают пищевые предпочтения животных и лишь частично, локальный растительный покров вокруг норок. Реконструкция сумм положительных температур показала, что палиноспектры отражают троекратные колебания условий вегетации: два кратковременных периода существенного ухудшения условий вегетации (сумма положительных температур около 200-300 град. × дней), около 46, 34-33 тыс. кал. лет. (и два периода улучшения условий вегетации 45 – 43 тыс. кал. лет (сумма положительных температур около 600 град. х дней) и 32 тыс. кал лет (сумма положительных температур около 750 град. х дней).
Ключевые слова:
повторно-жильные льды, поздний плейстоцен, многолетнемерзлые породы, едома, пыльца, споры, радиоуглерод, Зеленый Мыс, низовья Колымы, северо-восток Якутии
Работа выполнена при финансовой поддержке РНФ (грант 19-17-00126).
Abstract: The pollen characteristics of the Zelyony Mys yedoma record cyclical changes in vegetation cover, both on a local and regional scale. There are three local pollen zones such as: Varia +Artemisia, Varia + Selaginella sibirica, and Pinus + Betula sect. Nanae. Pollen spectra reflect open landscapes of mosaic hypo-arctic tundra. Some local pollen zones in the ice wedges correspond to the local pollen zones of yedoma: Varia + Selaginella sibirica dates from 41.9-33.8 CalBP, Pinus + Betula sect. Nanae (P+B) is approximately dated to 33.5-30 CalBP. Reconstruction of the annual thawing index showed several fluctuations: two short-term periods of significant deterioration of vegetation conditions (annual thawing index was about 200-300 deg. × days, about 46, 34-33 CalBP), and two periods of improvement of vegetation conditions 45-43 and 32 CalBP (annual thawing index was about 600-750 deg. x days). Regional pollen in burrows is the same in synchronous horizons of yedoma. Noticeable amount of larch pollen (10%) in the fossil burrows in the Zelyony Mys yedoma showed that favorable conditions are created in the burrows for the preservation of larch pollen. The seeds found in minks reflect animal dietary preferences and only partlially local vegetation around minks. Reconstruction of positive temperature sums indicates the palinospectra reflect three-fold fluctuations of vegetation conditions: two short periods of substantial deterioration of vegetation conditions (the sum of positive temperatures is about 200-300 degrees x days), about 46, 34-33 thousand years (and two periods of improvement of vegetation conditions 45-43 thousand years (sum of positive temperatures about 600 degrees x days) and 32 thousand years (sum of positive temperatures about 750 degrees x days).
Keywords: Ice wedge, Late Pleistocene, permafrost, yedoma, pollen, spores, radiocarbon, Zelyony Mys exposure, Lower Kolyma River, north-eastern Yakutia
ВВЕДЕНИЕ
Сингенетические повторно-жильных льды и едомные отложения Зеленого Мыса авторами ранее детально описаны [1,2], однако палинологическим особенностям этого разреза уделено недостаточное внимание.
Цель работы авторов: выполнить палинологический анализ повторно-жильных льдов и едомных отложений Зеленого Мыса и органического материала из норок сусликов (рис. 1, 2).
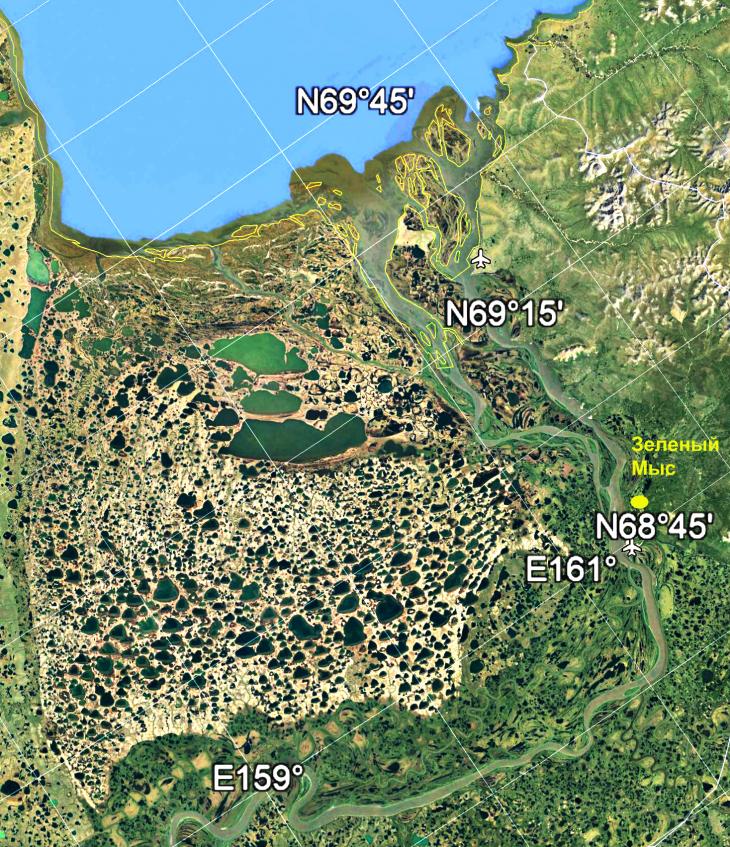
Рис. 1. Расположение разреза Зеленый Мыс в низовьях Колымы. Основа карты взята с Google map

Рис. 2. Расположение разреза Зеленый Мыс в низовьях Колымы (на переднем плане - аэропорт, за ним поселок Черский, на заднем плане - порт Зеленого Мыса)
СТРОЕНИЕ И СОСТАВ ИССЛЕДОВАННЫХ ЕДОМНЫХ ТОЛЩ
Расположение и строение едомной толщи
Едомная толща Зеленого Мыса вскрывалась с 1983 г. по 2006 гг. в овраге в 2 км севернее пос. Зеленый Мыс на правом берегу р.Колымы (рис. 3). Среди едомных толщ Нижнеколымского региона -это одно из самых представительных обнажений, в наиболее полном виде его вертикальная стенка достигала 36 м (рис. 4).

Рис. 3. Едомная толща Зеленого Мыса, характер обнажённости, которой 1 октября 1983 г. послужил триггером Ю.Васильчуку [Васильчук, 1992, 1999] для формулирования гипотезы мезо- и макроциклитности сингенетических повторно-жильных льдов, I-III ярусы – горизонты, насыщенные органическим материалом – остатками мхов, трав, корешками, стебельками, мелкими веточками, накапливавшиеся в субаэральном режиме

Рис. 4. Мощная едомная толща Зеленого Мыса, вскрытая в 1987 г, высота обнажения 36 м.
Обнажение высотой около 36 м состоит из двух частей: верхняя серая мощностью около 10 м, практически без органического материала, и нижняя - коричневато серая, в которой переслаиваются оторфованные пачки с растительным детритом.
Отложения повторно-жильного комплекса, представлены темно-серыми и коричневато-серыми супесями. Мощные повторно-жильные льды пронизывают всю 36-метровую толщу. В нижней части жилы несколько более массивны (рис. 4), шире (до 2–2,5 м во фронтальном разрезе), вверх нередко сужаются (до 0,5–1 м), но зато располагаются чаще (расстояние между жилами уменьшается – до 2–4 м). У крупных жил на уровне торфяных горизонтов нередко выражены плечики.
Климат и растительность
Территория нижнего течения р.Колымы относится к арктическому и субарктическому климатическим поясам континентальной области [3]. Безморозный период составляет в среднем 55 дней, сумма положительных температур − 800–900 °С. Средняя температура января −34 °С, средняя температура июля +9 °С [4]. Годовая сумма осадков составляет 150 мм, большая часть осадков выпадает в виде снега [5]. Зональная растительность в районе разреза Зеленый Мыс представлена лиственничными редколесьями (рис. 5), лесообразующая порода лиственница Каяндера (Larix cajanderi Mayr.)

Рис. 5. Растительность в районе разреза Зеленый Мыс. Вид с горы Родинка в направлении поселка Черский. Фото П.Крестова
Кустарниковый ярус образуют: березка тощая (Betula exilis Sukacz.), ива сизая (Salix glauca L.), шиповник иглистый (Rosa acicularis Lindl.), встречается кедровый стланик (рис. 5). Кустарнички в большинстве относятся к представителям семейства верескоцветных: голубика болотная (Vaccinium uliginosum L.), арктоус красноплодный (Arctous erytrocarpa Small.), и альпийский A. alpina (L.) Niedenzu. Разнотравье представлено семействами гречишных Bistorta vivipara (L.) Gray, сложноцветных Antennaria dioica (L.) Gaertn., Artemisia arctica Less., розоцветных Potentilla stipularis L. и др. [6]. Непосредственно у разреза Зеленый мыс травянистый ярус образован в основном представителями семейств кипрейных - иван-чаем (Chamaenerion angustifolium (L.) Scop.), сложноцветных пижмой (Tanacetum vulgare L.), мелколепестником (Erigeron acris L.), злаков – мятликом (Poa pratensis L.), ячменем (Hordeum jubatum (L.) Nevski), в кустарниковом ярусе ива сизая (Salix glauca L.) [7].
Методы и результаты
Радиоуглеродное датирование. По данному разрезу получено 14 конечных радиоуглеродных датировок (рис. 6, 7) в том числе по мамонтовым останкам (рис. 8) и из нижней части разреза 5 датировок, выходящих за пределы датирования радиоуглеродным методом.
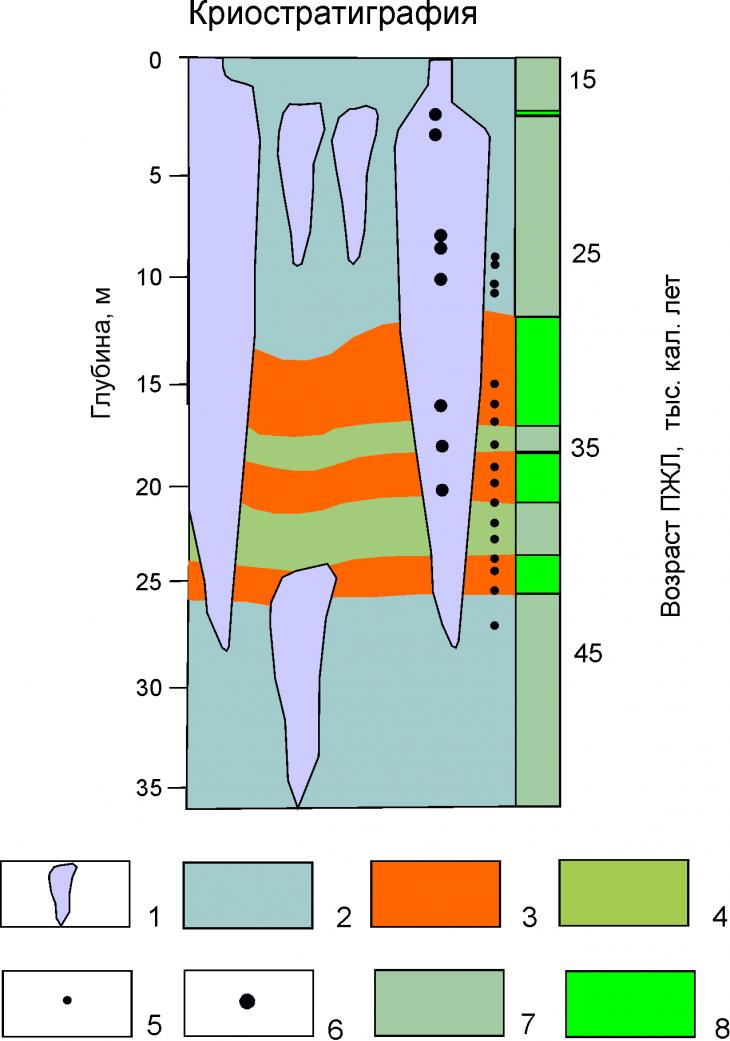
Рис. 6. Разрез едомной толщи Зеленого Мыса с калиброванными 14С датировками, из [2]:
1 – лед повторно-жильный; 2 – супесь серая, мерзлая со среднеслоистой и сетчатой, средне– и тонкошлировой криотекстурой; 3 – торф мерзлый с косослоистыми, решетчатыми криотекстурами; 4 – супесь коричнево-серая, мерзлая со тонкослоистой, средне– и тонкошлировой криотекстурой; отбор образцов для анализа пыльцы и спор: 5 – из вмещающих отложений, 6 – изо льда; 7 – отложения субаквального этапа накопления едомы; 8 – отложения субаэрального этапа накопления едомы
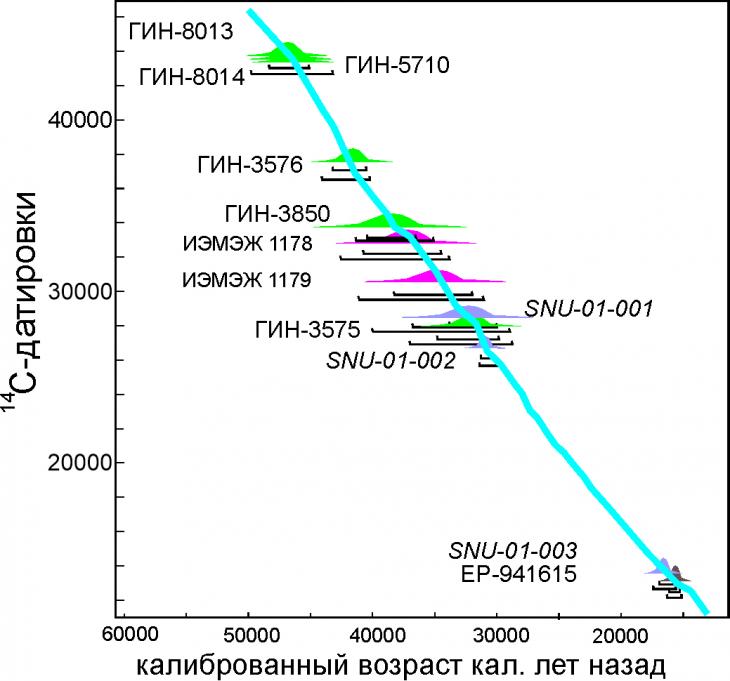
Рис. 7. Калиброванные 14С датировки едомной толщи Зеленого Мыса[2]

Рис. 8. Мамонтовая берцовая кость в разрезе едомы Зеленый Мыс, с глубины 12 м, датированная 43700 ±800 лет (ГИН-3849). Фото Л.Д.Сулержицкого
Согласно полученным датировкам [2] едома Зеленого мыса накапливалась в течение 33 тыс. кал. лет - интервале 48-15 тыс. кал. лет.
МЕТОДЫ ПОЛЕВЫХ И ЛАБОРАТОРНЫХ ОПРЕДЕЛЕНИЙ
Палинологический анализ повторно-жильных льдов и вмещающих отложений. Спорово-пыльцевая анализ выполнен А.К.Васильчук, палинологические диаграммы построены, как по образцам из вмещающих отложений, так и отобранным непосредственно из ПЖЛ. Расчет компонентов производился от суммы пыльцы и спор. Проанализировано 56 образцов, из них 45 из вмещающих отложений и 11 из повторно-жильных льдов. Чтобы исключить загрязнение образца современной пыльцой и спорами, каждый образец промывался после частичного таяния талой водой. Каждый образец льда был помещен в 300-мл полиэтиленовый флакон высокой плотности после полного таяния и оставлен на 24 часа перед разделением на пластиковые флаконы для различных анализов. Остаток со дна флакона отбирался для анализа пыльцы с целью получения достаточной концентрации пыльцы и спор. Обработка образцов из отложений едомы производилась с использованием сепарационного метода, разработанного В.П. Гричуком [8].Обработка образцов льда для анализа пыльцы и спор включала испарение остатка, дефлокуляцию с использованием KOH, центрифугирование, фильтрацию через сита 40 мкм, 10 мкм и 2 мкм и помещение образцов пыльцы в глицерин. Идентификация пыльцы и спор под световым микроскопом при увеличении 400х с использованием авторской справочной коллекции лаборатории геоэкологии Севера МГУ им. М. В. Ломоносова, российской базы данных пыльцы (pollendata.org.) и определителей [9, 10]. Пыльца березы была разделена на Betula sect Albae) и Betula sect. Nanae [9]. Пыльца сосны была идентифицирована как пыльца сосны обыкновенной (Pinussylvestris), сосны сибирской (Pinussibirica) и кедрового стланика (Pinispumila) на основе авторской коллекции поверхностных образцов, авторской справочной коллекции пыльцы, а также описаний пыльцы [10]. Из-за низкой концентрации пыльцы и спор во льду подсчитывались все микрофоссилии в полученном концентрате во фракции 10-40 мкм. Фракция 2-10 мкм использовалась для определения содержания мелких спор зеленых мхов и спор грибов. Концентрация рассчитывалась как отношение подсчитанных пыльцевых зерен к объему пробы. Очевидно, что полученные значения концентрации отличаются от тех, которые можно было бы получить с использованием таблеток Lycopodiumclavatum.
Сумма положительных температур рассчитана согласно анализу субфоссильных палиноспектров [11]. Палиноспектры Арктики разделяются на основные широтно-температурные группы: 1) арктических тундр и полярных пустынь, где сумма положительных температур ниже 400 градусо×дней; 2) северных гипоарктических тундр, где сумма положительных температур от 500 до 700 градусо×-дней; 3) южных гипоарктических тундр, где сумма положительных температур от 700 до 800-900 градусо×дней; 4) подзоны крупных стлаников, лесотундры (и частично северной тайги), где сумма положительных температур 800-1200 градусо×дней. Эти группы, в целом имеют широтное простирание, они достаточно явно выделяются по типам растительности и относительно контрастны по пыльцевым характеристикам. Для поздненеоплейстоценовых ландшафтов севера Сибири определяющими своеобразие факторами помимо температурного режима были высота снежного покрова и связанная с ней влажность почв [12]. Это во многом определило своеобразие позднеплейстоценовой растительности. Применение данной методики реконструкции суммы положительных температур дало возможность определить относительные изменения палиноспектров в отдельных образцах и выразить их в единых терминах суммы положительных температур. Предложенная интерпретация позволяет сопоставлять между собой датированные спорово-пыльцевые диаграммы разрезов, расположенных на значительном удалении друг от друга. Ограничением данного метода является его точность, что связано с воздействием на процесс формирования палиноспектров тафономических факторов.
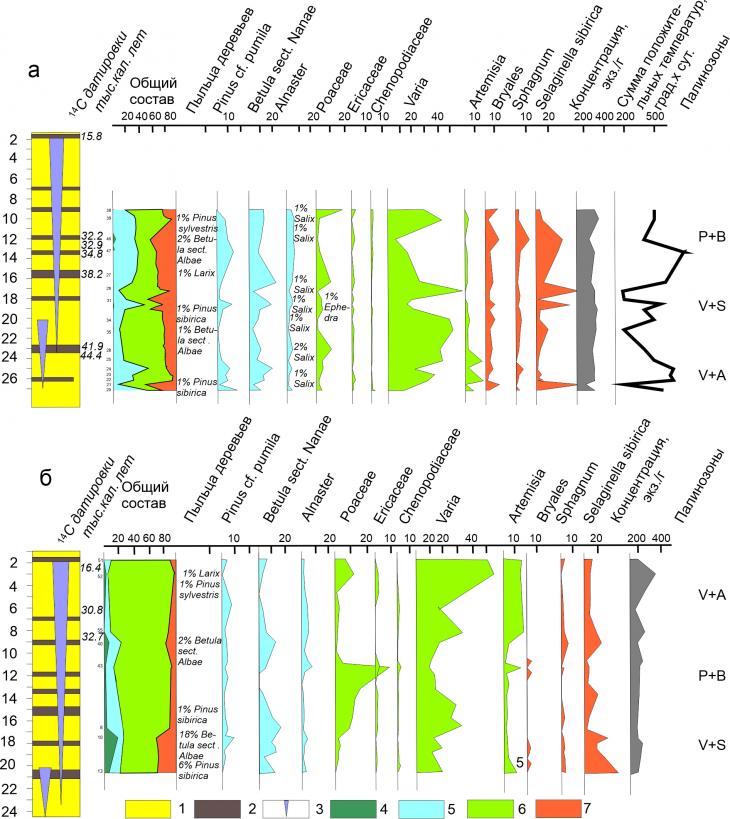
Рис. 9. Спорово-пыльцевая диаграмма по результатам анализа отложений едомы (а) и повторно-жильных льдов (б): 1- супесь, 2 – торф, 3 – повторно-жильный лед; процентное содержание: 4 – пыльцы древесных пород, 5 – пыльцы кустарников, 6 – пыльцы трав, 7 – спор.
РЕЗУЛЬТАТЫ АНАЛИТИЧЕСКИХ ОПРЕДЕЛЕНИЙ
Анализ совокупности 14С датировок показал, что период формирования едомных толщ Зеленого Мыса составил около 33 тыс. лет от 48 до 15 кал. тыс. лет. Полученная спорово-пыльцевая диаграмма из вмещающих отложений датируется примерно в интервале от 30 до 46 тыс. кал. лет назад (рис. 9, а). В интервале глубин, 9.5-27.0 м, который охарактеризован палинологически, выделены три цикла смены субаквальных условий субаэральными. Скорость накопления была неравномерной в течение субаквальных фаз была существенно выше, чем в течение субаэральных фаз и варьировала от 0.2 до 1.0 м за тысячу лет. Всего в разрезе отмечено 4 субаэральные и 4 субаквальные фазы [2]. Субаэральные фазы были продолжительнее субаквальных, по результатам 14С датирования они длились по 2–4 тыс. лет. Субаквальные фазы по длительности едва ли заметно превышали 1–1.5 тыс. лет. В разрезе наоборот, отложения, относящиеся к субаквальным фазам имеют гораздо большую мощность. Спорово-пыльцевая диаграмма из повторно-жильных льдов на основании радиоуглеродных датировок непосредственно из повторно-жильного льда и вмещающих отложений [2] датируется нами в интервале 16–46 тыс. кал. лет назад (рис. 9, б).
Палинологическая характеристика повторно-жильных льдов и вмещающих отложений
Палинологическая характеристика разреза едомных отложений Зеленый Мыс дает основание для выделения на диаграмме трех локальных палинозон: Varia +Artemisia (V+A), (интервал 27.0–24 м) – Varia + Selaginella sibirica (V+S) (интервал 24–18 м) и Pinus + Betula sect. Nanae (P+B), (интервал18.0–9.5 м). Практически все палиноспектры отражают открытые ландшафты мозаичных гипоарктических тундр.
Палинозона V+A датируется примерно 46-41.9 тыс. кал. лет. назад. Максимум недоразвитой пыльцы разнотравья отмечен на глубине 26.6 м и локальный максимум (4%) на глубине 24.5 м. В пределах палинозоны выделяется локальный максимум содержания пыльцы кедрового стланика. Распределение пыльцы кедрового стланика аналогично распределению пыльцы карликовой березки, что отражает и усиление ветрового переноса, и улучшение условий вегетации в региональном масштабе. Снижению содержания пыльцы кедрового стланика соответствует повышение содержания пыльцы полыни скорее всего это означает изменение режима увлажнения. Во всех образцах присутствует пыльца ольховника. Среди трав отмечена пыльца Angelica, Aster, Brassicaceae, Koenigia, Polygonum viviparum, Rosaceae, Potentilla-type, Rumex/Oxyria digyna type, Saxifragaceae, Onagraceae и др., т.е. по составу это в основном арктическое разнотравье.
Палинозона V+S (24–18 м), приблизительно датируется в интервале 41,9–33,8 тыс. кал. лет назад и характеризуется повышенным содержанием недоразвитой пыльцы двудольных. Практически вся пыльца разнотравья за редким исключением недоразвита. Пыльца древесных встречается единично. Зато отмечаются локальные максимумы карликовой березки и ольховника. Отмечена пыльца Caryophyllaceae, в том числе и Silene-type, а также пыльца Draba, Saxifragaceae и Ephedra. Палиноспектры отражают ландшафты, формировавшиеся при коротком вегетационном периоде но с достаточно высокими температурами.
Палинозона P+B (18–9,5 м) приблизительно датируется 33.5–30 тыс. кал. лет назад. Эта палинозона характеризуется резкими колебаниями содержания основных компонентов, таких как пыльца разнотравья (Varia) и спор Selaginella sibirica и Betula sect. Nanae. На фоне резких колебаний содержания этих региональных компонентов содержание пыльцы кедрового стланика изменяется плавно, демонстрируя скорее дальний занос.
Двум из этих палинозон соответствуют палинозоны выделенные по спорово-пыльцевым спектрам непосредственно из ПЖЛ (естественно со смещенным положением по вертикали вниз).
Палинозона V+S выделена на глубине 20–16 м, она вероятно соответствует палинозоне V+S, выделенной на диаграмме вмещающих отложений, что не противоречит возрасту 41,9–33,8 тыс. кал. лет. Палиноспектры характеризуются повышенным содержанием недоразвитой пыльцы двудольных среди трав встречена пыльца Caryophyllaceae, в том числе и Silene-type,а также пыльца Draba, Saxifragaceae и Ephedra. Содержание пыльцы древесных здесь максимальное 20%, в основном это пыльца березы. Повышенное содержание пыльцы березы во льду отражает пыльцу из более южных районов, выпавшую на поверхность снега.
Палинозона P+B (16–9,5 м) приблизительно датируется 33.5–30 тыс. кал. лет назад. Эта палинозона характеризуется пиковым содержанием пыльцы злаков (до 48%), при этом с пиком содержания пыльцы злаков совпадают локальные пики полыни и ольховника, а также резкое снижение содержания пыльцы карликовой березки Пыльца хвойных встречается единично.
Палинозона V+A датируется примерно 30-16 тыс. кал. лет. назад. Максимум недоразвитой пыльцы разнотравья отмечены на глубине 26.6 м и локальный максимум (4%) на глубине 24.5 м. В пределах палинозоны выделяется максимальное содержание недоразвитой пыльцы разнотравья и локальный максимум содержания пыльцы злаков, а также заметное содержание пыльцы полыни. Среди трав отмечена пыльца Brassicaceae, Koenigia, Polygonum viviparum, Rosaceae, Potentilla-type, Rumex/Oxyria digyna type, Saxifragaceae, и др., т.е. по составу это в основном арктическое разнотравье.
Спорово-пыльцевые спектры из норок сусликов
Палиноспектры из гнездовых и кормовых норок в разрезе Зеленый Мыс (табл. 1) дают представление о составе регионального пыльцевого дождя и о локальных биоценозах. Региональный пыльцевой дождь представлен пыльцой деревьев и кустарников (березы, карликовой березки и ольховника) , а также ветроопыляемых растений (злаков, осок и полыней). В норке Р-1225Е идентифицирована в заметном количестве пыльца лиственницы (10%), имеющая очень хорошую сохранность. Скорее всего лиственница произрастала в окрестностях норки, поскольку в поверхностных пробах в лиственничных редколесьях в низовьях Колымы содержание пыльцы лиственницы не превышает 2-5% [11]. Очевидно, что условия для сохранности пыльцы лиственницы в норках лучше, чем в отложениях ледового комплекса. Важно отметить, что в палиноспектрах норок встречается пыльца трав семейств Polygonaceae, Rosaceae, Caryophyllaceae, макроостатки которых, а также их семена также обнаружены в норках.
Таблица 1. Процентное содержание пыльцы и спор из норок грызунов в сингенетических едомных поздненеоплейстоценовых отложениях в разрезе Зеленый Мыс, правобережье в низовьях р. Колымы
|
пыльца и споры
|
номер образца
|
|
р-1208
|
р-1209**
|
Р-1225Е
|
97/3, помет мыши***
|
|
содержание пыльцы и спор, %
|
|
Пыльца деревьев
|
2.5
|
2.2
|
10.0
|
4.9
|
|
Пыльца кустарников
|
21.5
|
33.0
|
2.5
|
26.4
|
|
Пыльца трав и кустарничков
|
50
|
48.3
|
38.7
|
36.3
|
|
Споры
|
26.2
|
16.5
|
48.8
|
32.4
|
|
Larix
|
–
|
–
|
10.0
|
–
|
|
Betula sect.Albae
|
2.5
|
2.2
|
–
|
4.9
|
|
Pinus pumila
|
1.2
|
3.3
|
–
|
–
|
|
Betula sect. Nanae
|
20
|
19.8
|
2.5
|
21.6
|
|
Alnaster
|
–
|
5.5
|
–
|
4.9
|
|
Salix
|
–
|
4.4
|
–
|
–
|
|
Poaceae (крупные)
|
16.2
|
1.1
|
7.5
|
6.8
|
|
Poaceae (мелкие)
|
5
|
4.4
|
–
|
10.7
|
|
Cyperaceae
|
11.2
|
14.3
|
–
|
8.8
|
|
Ericaceae
|
5.0
|
8.8
|
–
|
2.0
|
|
Caryophyllaceae
|
2.5
|
2.2
|
8.7
|
–
|
|
Lamiaceae
|
–
|
–
|
–
|
–
|
|
Polemoniaceae
|
–
|
–
|
–
|
2.0
|
|
Draba sp.
|
–
|
–
|
1.3
|
9.8
|
|
Rumex sp
|
–
|
–
|
5.0
|
2.0
|
|
Sparganium sp
|
–
|
–
|
1.3
|
–
|
|
Oxiria digina
|
–
|
–
|
–
|
1.0
|
|
Potentilla sp.
|
–
|
–
|
2.5
|
4.0
|
|
Liliaceae
|
–
|
4.4
|
–
|
–
|
|
Polygonum cf. aviculare
|
–
|
1.1
|
–
|
–
|
|
Polygonum cf. bistorta
|
–
|
1.1
|
–
|
–
|
|
Chenopodiaceae
|
2.5
|
–
|
–
|
–
|
|
Artemisia
|
2.5
|
5.5
|
–
|
–
|
|
Varia
|
5.0
|
11.1
|
12.5
|
–
|
|
Bryales
|
18.8
|
8.8
|
17.0
|
16.7
|
|
Sphagnum sp.
|
3.7
|
1.1
|
10.0
|
–
|
|
Polypodiaceae
|
–
|
–
|
–
|
–
|
|
Selaginella sibirica
|
3.7
|
6.6
|
20.0
|
–
|
|
Equisetum sp.
|
–
|
–
|
|
15.7
|
|
Lycopodium sp(cf.clavatum)
|
–
|
–
|
1.3
|
–
|
|
*переотлож.
|
7.5
|
2.1
|
1
|
–
|
|
Экз.
|
80
|
91
|
80
|
102
|
*переотложенные доплейстоценовые палиноморфы
** споры грибов 10 экз., уголь менее 10 мкм – 115 экз.
***споры грибов 3 экз.
Сборы С.В. Губина и О.Г. Заниной, палинологический анализ А.К.Васильчук
Дискуссия
Судя по совокупности 14С датировок общий период формирования едомных толщ Зеленого Мыса длился около 33 тыс. лет от 48 до 15 кал. тыс. лет. Нижний предел формирования едомной толщи зафиксирован тремя датировками по крупным веткам и костям мамонтовой фауны, а верхний предел определен 14С датировкой из погребенной почвы, отобранной А.Пфеффер близ кровли ММП 15.7 кал тыс. лет и AMS датировкой, полученной непосредственно из ПЖЛ - 16.4 кал тыс. лет. [2].
Для корректного датирования также очень важны данные по запасам семян в ископаемых норках сусликов, поскольку датировки по ним получены по заведомо синхронному органическому материалу [13]. Норы сусликов указывают на субаэральную фазу развития полигонального массива, когда ледяные жилы активно росли в ширину. Подобные находки, свидетельствуют в циклическом переменном субаквально-субаэральном генезисе сингенетических полигонально-жильных комплексов.
П.Баккер с соавторами [14] комбинируя моделирование летних температур во время гляциального максимума (LGM) из второй и третьей фазы Проекта сопоставления моделирования палеоклимата (Paleoclimate Modelling Intercomparison Project, 2, 3) с экспериментами чувствительности факторов с использованием модели CESM (Community Earth System Model) пришли к выводу, что локальные летние температурные аномалии, полученные в результате реконструкций на севере Сибири, пока не объяснимы с точки этих моделей. В других районах реконструкция летних температур совпадает с результатами полевых исследований. Т.е. для севера Евразии очевидно не учитываются какие-то факторы, которые для более южных регионов существенны. Отметим, что Н.Вайцель с коллегами [15] привели свою версию реконструкции палеоклимата Сибири по палинологическим данным. Основной результат исследования заключается в том, что для территории Сибири в период гляциального максимума (LGM) по результатам палинологических исследований летние температуры выше, чем по результатам моделирования в проекте PMIP3 (Paleoclimate Modelling Intercomparison Project - PMIP3). Очень показательны выполненные В. Мейером с соавторами палеореконструкции летних температур на Камчатке и в сопредельных регионах [16] для периода последних 20 тыс. лет. По результатам интерпретации летние температуры на Камчатке в период от 20 до 18 тыс. кал. лет назад были близки к температурам в позднем голоцене. Около 18 тыс. кал. лет назад летние температуры резко снизились и оставались низкими до 14.6 тыс. кал. лет назад, но после этого периода летние температуры вернулись к «голоценовым» значениям. Поэтому палеотемпературные реконструкции важны для адекватной оценки особенностей летнего температурного режима в Северной Якутии в позднем неоплейстоцене.
Результаты палинологического исследования (рис. 9 а, б) показали, что условия осадконакопления практически во время всего периода формирования отложений были благоприятными для накопления не только локальных, но и региональных компонентов. Максимумы содержания пыльцы разнотравья совпадают с минимумами содержания пыльцы кедрового стланика. Отметим, что пыльца кедрового стланика здесь крупнее, чем пыльца современного кедрового стланика, отобранная авторами в районе пос. Ваеги на юго-западе Чукотки. Пыльца разнотравья представлена семействами розоцветных, камнеломковых, лютиковых. Семена растений из этих семейств (PotentillaniveaL., DrabacinereaAdam., Ranunculusrepens L.), были обнаружены разрезе Зеленый Мыс в норках сусликов в интервале глубин от 5 до 15 м [17], что дополняет результаты спорово-пыльцевого анализа. Одновременная реакция и локальных и региональных составляющих спорово-пыльцевых спектров означает, что изменения условий вегетации были обусловлены региональными факторами и связаны с определенными климатическими колебаниями условий периода вегетации, а не изменениями локальных условий осадконакопления. Максимумы содержания пыльцы сибирского кедра и спор приурочены к горизонтам, находящимся непосредственно под торфяниками, тогда как в самих торфяных горизонтах весьма высоко содержание пыльцы кустарников (в основном березы карликовой – до 26%), хотя макроостатки кустарников в торфяных прослоях этого разреза не встречены.
Реконструкция сумм положительных температур показала, что палиноспектры отражают троекратные колебания условий вегетации. два кратковременных периода существенного ухудшения условий вегетации (сумма положительных температур около 200-300 град. × дней), около 46, 34-33 тыс. кал. лет назад (и два периода улучшения условий вегетации 45 – 43 тыс. кал. лет (сумма положительных температур около 600 град.х дней) и 32 тыс. кал. лет назад (сумма положительных температур около 750 град.х дней).
Спорово-пыльцевые спектры палинозоны V+S (26–22 м), отвечают ландшафтам арктических тундр с коротким вегетационным сезоном, выделяется два цикла длительностью 1,5-2 тыс.лет Сумма положительных температур колеблется от 200 до 400 град. × дней, при коротком вегетационном периоде. Короткий вегетационный период означает, что температура вегетационного периода была достаточно высокой, выше, чем в современных арктических тундрах, но его длительность не давала возможности даже для вызревания пыльцы разнотравья. Спорово-пыльцевые спектры палинозоны P+B (20–9,5 м) Выделяется три периода снижения суммы положительных температур и три периода их повышения. Около 34 – 33 тыс. кал. лет назад (глубина 18 м), сумма положительных температур составляла приблизительно 400 град. х дней. Здесь споры Selaginella sibirica являются доминантами палиноспектров. Около 33-32 тыс. лет назад отмечается повышение суммы положительных температур до 650 град х дней. Отмечен пик содержания пыльцы сосны (22%), и одновременный максимум пыльцы Betula sect. Nanae (20%). Недоразвитая пыльца разнотравья почти отсутствует, что говорит о достаточной для вызревания пыльцы длительности вегетационного периода. Приблизительно в интервале 33–31 тыс. кал. лет назад увеличилась скорость осадконакопления, за это время накопились отложения на глубине 18,1–17,2 м. В это время условия сезона вегетации снова ухудшились: сумма положительных температур составляла около 150–200 град. × дней, при коротком вегетационном сезоне, об этом свидетельствует доминирование недоразвитой пыльцы разнотравья и спор селагинеллы. Около 31–30 тыс. лет назад, что соответствует отложениям на глубине 13,6–16,4 м, отмечены максимум содержания пыльцы Betula sect. Nanae (26%), который сменяет максимум пыльцы Pinus sylvestris (22%). Для севера Евразии этот период улучшения условий вегетации прослеживается наиболее определенно и его можно выделить почти на всех спорово-пыльцевых диаграммах. Палиноспектры, полученные из повторно-жильных льдов, подтверждают предположение об их региональном характере. Здесь содержание пыльцы древесных пород и пыльцы полыни заметно выше. Палиноспектры из повторно-жильных льдов отражают ритмические колебания состава регионального весеннего пыльцевого дождя.
Палинологическая характеристика отложений разреза Зеленый Мыс фиксирует примерно семь среднепериодных циклических изменений растительного покрова, как в локальном, так и в региональном масштабе. Это выразилось в резкой смене доминантов: палиноспектры с преобладанием пыльцы карликовой березки с участием пыльцы сосны обыкновенной, сменяются палиноспектрами с доминированием пыльцы злаков и полыни или с доминированием недоразвитой пыльцы разнотравья и спор плаунка сибирского. При этом в пределах выделенных палинозон преимущественно сохраняется характер циркуляции атмосферы в весенне-летний период, циклы отражают изменения базиса эрозии и колебания температур вегетационного периода, а также его длительность.
Ископаемые норы сусликов, единичные или их фрагменты, на территории Нижнеколымской низменности были встречены Т.Н.Каплиной, С.В.Губиным и др. в обнажениях Дуванный Яр на реке Колыма, Станчиковский Яр на реке Анюй, ряде озерных отложений в бассейне реки Большая Чукочья. В восточной части разреза Воронцовский Яр В.К.Рябчун обнаружил большую кладовую суслика на глубине 12 м, из которой В.П.Никитиным были сделаны определения макроостатков и семян растений. В норе были собраны в основном коробочки ясколки (Cerastiumsp.) и плоды лапчатки (Potentillamultifida, Potentilla sp.), в меньшем количестве отмечены стручки и семена крупки (Draba sp.) и семена полярного мака [18].
Сотрудники Пущинского института биофизики клетки и Института физико-химических и биологических проблем почвоведения сумели вырастить смолёвку (Silene stenophylla) - растение из семейства гвоздичных из семян, отобранных из норки суслика в мерзлой толще Дуванноярской едомы (семена датированы по радиоуглероду 36.1 тыс. кал. лет (Beta-157195) [19]. А.В. Шатилович с соавторами [20] удалось выделить из материала погребённой норы суслика на глубине около 30 м в мёрзлой стенке обнажения Дуванный яр жизнеспособные многоклеточные организмы – почвенные нематоды (круглые черви). Из подобных нор здесь, датированы растительные остатки около 32 тыс. лет [21]. Нематоды были обнаружены также в образце едомных многолетнемёрзлых отложений при колонковом бурении в районе р. Алазея с глубины 3,5 м содержавшем слаборазложившиеся растительные остатки (датированных по радиоуглероду 44.7 тыс. кал. лет, AA-109003). . Таким образом, впервые были получены данные, свидетельствующие о способности многоклеточных животных переживать длительный (десятки тысяч лет) криобиоз в условиях естественной криоконсервации. Органический материал из норок млекопитающих – один из лучших объектов для датирования в криолитозоне из-за того, что обитатели норок привносят туда только современную им органику. Гнездовые норы малого суслика располагаются на незаливаемых во время весеннего снеготаяния повышениях микрорельефа. На территориях, периодически затопляемых водой, не возникает устойчивых поселений млекопитающих. Поэтому органический материал в норках как правило автохтонный и радиоуглеродные датировки по этому материалу надежны
В овраге у пос. Зеленый Мыс на расстоянии 50 м С.В. Губиным было встречено шесть ископаемых нор сусликов. Их фрагменты располагались на глубине от 5 до 15 м от дневной поверхности. Норы были изучены в 100-200 м от обнажения, детально исследованного авторами в 1983-1987 гг. (см. рис. 3 и 4). В норе на глубине 14 м от дневной поверхности сохранился сантиметровый слой семян: луковички (клубнепочки) змеевика живородящего (горца живородящего) Bistorta vivipara (L.) S.F. Gray (Poligonum viviparum L.), здесь также обнаружены семена крупки серой Draba cinerea Adam и мало потемневшие семена лютика ползучего Ranunculus repens L. [17]. В соседней норке обнаружены семена осок (Саrех), мятлика (Роа), лапчатки двух видов, в том числе лапчатки снежной Potentilla nivea L., лютика ползучего Ranunculus repens L., крупки серой Draba cinerea Adam., смолевки узколистной Silene stenophylla Ledeb., а также луковички змеевика живородящего Bistorta vivipara (L.) S.F.Gray. В нижняя камера норки на глубине 12 м от дневной поверхности была полностью заполнена семенами лапчатки снежной Potentilla nivea L., и лютика ползучего Ranunculus repens L. крупки серой Draba cinerea Adam., осоки прямостоящей Carex stans Drej. и смолевки узколистной Silene stenophylla Ledeb. Масса сухих семян в камере достигла 650 г, что в пересчете на отдельные экземпляры составило 845 тыс. штук. 14С-датированием материала семян из рассматриваемой норы был определен их возраст, который составил 37.8 тыс.кал.лет (ИЭМЭЖ-1178) [17]. Анализ образа жизни современного длиннохвостого суслика (Citellus undulatus Pall.), населяющего арктические и субарктические районы северо-востока Евразии, показывает, что заготовку растительного корма зверек проводит в радиусе первых сотен метров от своей норы, а запасы семян и плодов могут достигать веса 6 кг и более. В то же время следует учитывать, что, расселяясь крупными колониями, эти животные существенным образом меняют облик поверхности, непосредственно прилегающей к норам. При этом резко усиливается дренаж участков, активизируются процессы гумификации и нитрификации органического материала, уничтожается исходный растительный покров, на смену которому в тундре приходят злаки и разнотравье [11]. Исследования современных нор показали, что семена, обнаруживаемые в норках, отражают пищевые предпочтения животных и лишь частично, локальный растительный покров. Норы сусликов нельзя считать надежным признаком существования ландшафтов степного типа, так как часто животные сооружают норки на аласах и гарях, в современной тайге в окрестностях Якутска [22]. Как показано Л.Г.Динесманом состав региональной составляющей палиноспектров в норках и синхронных им почвенных и торфяных горизонтах практически не отличается, а пыльца трав отражает зоогенные биоценозы, формирующиеся вокруг норок. Локальные компоненты отражают состав растительного покрова вокруг норок.
Это подтвердили палинологические исследования норок на разрезе Зеленый мыс. Палинологическая характеристика локальной составляющей почти во всех норках более постоянна, по сравнению с составом регионального пыльцевого дождя. Палиноспектры из норок характеризуют слабо характеризуют локальные фитоценозы и свидетельствуют об избирательной сохранности пыльцы и спор отдельных таксонов.
Состав регионального пыльцевого дождя по данным палинологического исследования норок ближе к палиноспектрам из повторно-жильных льдов, чем к палиноспектрам из вмещающих повторно-жильные льды отложений.
Выводы
1. Палинологическая характеристика разреза едомных отложений Зеленый Мыс дает основание для выделения на диаграмме трех локальных палинозон: Varia + Artemisia (V+A), (интервал 27.0–24 м) – Varia + Selaginella sibirica (V+S) (интервал 24–18 м) и Pinus + Betula sect. Nanae (P+B), (интервал18.0–9.5 м). Практически все палиноспектры отражают открытые ландшафты мозаичных гипоарктических тундр.
2. Палинозоны, выделенные по результатам палинологического повторно-жильных льдов, соответствуют палинозонам вмещающих льды отложений: Varia + Selaginella sibirica (V+S) выделена на глубине 20–16 м, что не противоречит возрасту 41,9–33,8 тыс. кал. лет. Палинозона Pinus + Betula sect. Nanae (P+B) в интервале 16–9,5 м приблизительно датируется 33.5–30 тыс. кал. лет назад.
3. Условия для сохранности пыльцы лиственницы в норках лучше, чем в отложениях ледового комплекса, это подтверждается высоким содержанием пыльцы лиственницы (10%) в одной из норок суслика в разрезе Зеленый Мыс.
4. Реконструкция сумм положительных температур показала, что палиноспектры отражают троекратные колебания условий вегетации: два кратковременных периода существенного ухудшения условий вегетации (сумма положительных температур около 200-300 град. × дней), около 46, 34-33 тыс. кал. лет. (и два периода улучшения условий вегетации 45–43 тыс. кал. лет (сумма положительных температур около 600 град. × дней) и 32 тыс. кал лет (сумма положительных температур около 750 град. × дней).
Библиография
1. Васильчук Ю.К., Есиков А.Д., Опруненко Ю.Ф., Васильчук А.К., Сулержицкий Л.Д. Новые данные по содержанию стабильных изотопов кислорода в сингенетических повторно-жильных льдах позднеплейстоценового возраста низовий р.Колымы // Доклады АН СССР. 1985. Том 281. N 4. С. 904–907
2. Васильчук Ю.К., Васильчук А.К. Реконструкция среднеянварской палеотемпературы воздуха 48-15 тысяч калиброванных лет назад по изотопно-кислородному составу едомы Зеленого Мыса // Криосфера Земли. 2021. Том XXV. №2. С. 48–61. doi: 10.15372/KZ20210205
3. Атлас СССР. М., 1986. 260 с.
4. Справочник по климату СССР. – Л., 1966. – Вып. 24, ч. 2. – 398 с.
5. Справочник по климату СССР. – Л., 1968. – Вып. 24, ч. 4. – 352 с.
6. Кожевников Ю.П. Ботанико-географические наблюдения на Колыме в районах среднего течения р. Березовка и поселка Черский // Биология и экология растений бассейна Колымы. Владивосток, ДВНЦ АН СССР, 1981, с. 99–117.
7. Занина О.Г., Лопатина Д.А. Возможности реконструкции состава растительных ассоциаций низовьев реки Колымы с помощью сопряженного (палинологического, фитолитного и карпологического) анализа // Криосфера Земли, 2017, т. XXI, № 3. С. 13–23.
8. Палеопалинология. Т. I. Ред. Покровская И.М. Л.: Недра, 1966. 352с.
9. Birks H.J.B. The identification of Betula nana pollen. New Phytologist 1968. Vol. 67(2). P. 309–314.
10. Бобров А.Е, Куприянова Л.А., Литвинцева Л.В., Тарасевич В.Ф. Споры папоротникообразных и пыльца голосеменных и однодольных растений флоры Европейской части СССР. Л., наука, 1983. 208 с.
11. Васильчук А.К. Особенности формирования палиноспектров в криолитозоне России, М.: Изд-во Моск ун-та, 2005, 245 с.
12. CAPE Project members. Holocene paleoclimate data from the Arctic: testing models of global climate change // Quaternaty Sciences Reviews. 2001. Vol. 20. P.1275–1287.
13. Zanina O. G. Fossil Rodent Burrows in Frozen Late Pleistocene Beds of the Kolyma Lowland // Entomological Review, 2005, Vol. 85, Suppl. 1. P. S133–S140.
14. Bakker P., Rogozhina I., Merkel U., Prange M. Hypersensitivity of glacial summer temperatures in Siberia // Climate of the Past, 2020, vol. 16. P. 371–386. doi: 10.5194/cp-16-371-2020.
15. Weitzel N., Hense A., Herzschuh U., Bohmer T., Cao X., Rehfeld K. A spatial reconstruction of Siberian Last Glacial Maximum climate from pollen data // Climate Dynamics, 2020. P. 1-25. doi: 10.31223/osf.io/76dfe
16. Meyer V.D., Hefter J., Lohmann G., Max L., Tiedemann R., Mollenhauer G. Summer temperature evolution on the Kamchatka Peninsula, Russian Far East, during the past 20 000 years // Climate of the Past, 2017, vol. 13, p. 359–377, doi:10.5194/cp-13-359-2017
17. Губин С.В., Максимович С.В., Занина О.Г. Анализ состава семян растений из ископаемых нор сусликов лессово-ледовых отложений обнажения Зеленый Мыс как показатель местных условий обитания // Криосфера Земли. 2001. Том 5. №2. С. 76–82.
18. Томирдиаро С.В. Лессово-ледовая формация Восточной Сибири в позднем плейстоцене и голоцене. Отв. ред. Н.И.Кригер. М.: Наука. 1980. 184 с.
19. Yashina S., Gubin S., Maksimovich S., Yashina A., Gakhova E., Gilichinsky D. Regeneration of whole fertile plants from 30,000-y-old fruit tissue buried in Siberian permafrost // Proceedings of the National Academy of Sciences (PNAS), 2012, vol. 109 (10). P. 4008–4013. doi: 10.1073/pnas.1118386109
20. Шатилович А.В., Чесунов А.В., Неретина Т.В., Грабарник И.П., Губин С.В., Вишнивецкая Т.А., Онстотт Т.С., Ривкина Е.М. Жизнеспособные нематоды из позднеплейстоценовых многолетнемёрзлых отложений Колымской низменности // Доклады Академии Наук, том 480, № 2. С. 253–255.
21. Губин С.В., Занина О.Г. Изменение почвенного покрова в ходе формирования отложений ледового комплекса на Колымской низменности (Часть 1) // Криосфера Земли, 2013, т. XVII, № 4. С. 48–56.
22. Динесман Л.Г. Изучение истории биогеоценозов по норам животных. М.: Наука. 1968. 100 с.
References
1. Vasil'chuk Yu.K., Esikov A.D., Oprunenko Yu.F., Vasil'chuk A.K., Sulerzhitskii L.D. Novye dannye po soderzhaniyu stabil'nykh izotopov kisloroda v singeneticheskikh povtorno-zhil'nykh l'dakh pozdnepleistotsenovogo vozrasta nizovii r.Kolymy // Doklady AN SSSR. 1985. Tom 281. N 4. S. 904–907
2. Vasil'chuk Yu.K., Vasil'chuk A.K. Rekonstruktsiya sredneyanvarskoi paleotemperatury vozdukha 48-15 tysyach kalibrovannykh let nazad po izotopno-kislorodnomu sostavu edomy Zelenogo Mysa // Kriosfera Zemli. 2021. Tom XXV. №2. S. 48–61. doi: 10.15372/KZ20210205
3. Atlas SSSR. M., 1986. 260 s.
4. Spravochnik po klimatu SSSR. – L., 1966. – Vyp. 24, ch. 2. – 398 s.
5. Spravochnik po klimatu SSSR. – L., 1968. – Vyp. 24, ch. 4. – 352 s.
6. Kozhevnikov Yu.P. Botaniko-geograficheskie nablyudeniya na Kolyme v raionakh srednego techeniya r. Berezovka i poselka Cherskii // Biologiya i ekologiya rastenii basseina Kolymy. Vladivostok, DVNTs AN SSSR, 1981, s. 99–117.
7. Zanina O.G., Lopatina D.A. Vozmozhnosti rekonstruktsii sostava rastitel'nykh assotsiatsii nizov'ev reki Kolymy s pomoshch'yu sopryazhennogo (palinologicheskogo, fitolitnogo i karpologicheskogo) analiza // Kriosfera Zemli, 2017, t. XXI, № 3. S. 13–23.
8. Paleopalinologiya. T. I. Red. Pokrovskaya I.M. L.: Nedra, 1966. 352s.
9. Birks H.J.B. The identification of Betula nana pollen. New Phytologist 1968. Vol. 67(2). P. 309–314.
10. Bobrov A.E, Kupriyanova L.A., Litvintseva L.V., Tarasevich V.F. Spory paporotnikoobraznykh i pyl'tsa golosemennykh i odnodol'nykh rastenii flory Evropeiskoi chasti SSSR. L., nauka, 1983. 208 s.
11. Vasil'chuk A.K. Osobennosti formirovaniya palinospektrov v kriolitozone Rossii, M.: Izd-vo Mosk un-ta, 2005, 245 s.
12. CAPE Project members. Holocene paleoclimate data from the Arctic: testing models of global climate change // Quaternaty Sciences Reviews. 2001. Vol. 20. P.1275–1287.
13. Zanina O. G. Fossil Rodent Burrows in Frozen Late Pleistocene Beds of the Kolyma Lowland // Entomological Review, 2005, Vol. 85, Suppl. 1. P. S133–S140.
14. Bakker P., Rogozhina I., Merkel U., Prange M. Hypersensitivity of glacial summer temperatures in Siberia // Climate of the Past, 2020, vol. 16. P. 371–386. doi: 10.5194/cp-16-371-2020.
15. Weitzel N., Hense A., Herzschuh U., Bohmer T., Cao X., Rehfeld K. A spatial reconstruction of Siberian Last Glacial Maximum climate from pollen data // Climate Dynamics, 2020. P. 1-25. doi: 10.31223/osf.io/76dfe
16. Meyer V.D., Hefter J., Lohmann G., Max L., Tiedemann R., Mollenhauer G. Summer temperature evolution on the Kamchatka Peninsula, Russian Far East, during the past 20 000 years // Climate of the Past, 2017, vol. 13, p. 359–377, doi:10.5194/cp-13-359-2017
17. Gubin S.V., Maksimovich S.V., Zanina O.G. Analiz sostava semyan rastenii iz iskopaemykh nor suslikov lessovo-ledovykh otlozhenii obnazheniya Zelenyi Mys kak pokazatel' mestnykh uslovii obitaniya // Kriosfera Zemli. 2001. Tom 5. №2. S. 76–82.
18. Tomirdiaro S.V. Lessovo-ledovaya formatsiya Vostochnoi Sibiri v pozdnem pleistotsene i golotsene. Otv. red. N.I.Kriger. M.: Nauka. 1980. 184 s.
19. Yashina S., Gubin S., Maksimovich S., Yashina A., Gakhova E., Gilichinsky D. Regeneration of whole fertile plants from 30,000-y-old fruit tissue buried in Siberian permafrost // Proceedings of the National Academy of Sciences (PNAS), 2012, vol. 109 (10). P. 4008–4013. doi: 10.1073/pnas.1118386109
20. Shatilovich A.V., Chesunov A.V., Neretina T.V., Grabarnik I.P., Gubin S.V., Vishnivetskaya T.A., Onstott T.S., Rivkina E.M. Zhiznesposobnye nematody iz pozdnepleistotsenovykh mnogoletnemerzlykh otlozhenii Kolymskoi nizmennosti // Doklady Akademii Nauk, tom 480, № 2. S. 253–255.
21. Gubin S.V., Zanina O.G. Izmenenie pochvennogo pokrova v khode formirovaniya otlozhenii ledovogo kompleksa na Kolymskoi nizmennosti (Chast' 1) // Kriosfera Zemli, 2013, t. XVII, № 4. S. 48–56.
22. Dinesman L.G. Izuchenie istorii biogeotsenozov po noram zhivotnykh. M.: Nauka. 1968. 100 s.
Результаты процедуры рецензирования статьи
В связи с политикой двойного слепого рецензирования личность рецензента не раскрывается.
Со списком рецензентов издательства можно ознакомиться здесь.
Рецензия на статью «Палинологическая характеристика едомы Зеленого Мыса и норок сусликов».
Предметом исследования являются палинологические особенности сингенетических повторно-жильных льдов и едомных отложений Зеленого Мыса в низовьях Колымы.
Основным методом исследования является спорово-пыльцевой анализ повторно-жильных льдов и вмещающих отложений. Палинологические диаграммы построены, как по образцам из вмещающих отложений, так и отобранным непосредственно из ПЖЛ. Расчет компонентов производился от суммы пыльцы и спор. Проанализировано 56 образцов, из них 45 из вмещающих отложений и 11 из повторно-жильных льдов. Чтобы исключить загрязнение образца современной пыльцой и спорами, каждый образец промывался после частичного таяния талой водой. Каждый образец льда был помещен в 300-мл полиэтиленовый флакон высокой плотности после полного таяния и оставлен на 24 часа перед разделением на пластиковые флаконы для различных анализов. Остаток со дна флакона отбирался для анализа пыльцы с целью получения достаточной концентрации пыльцы и спор. Обработка образцов из отложений едомы производилась с использованием сепарационного метода, разработанного В.П. Гричуком. Обработка образцов льда для анализа пыльцы и спор включала испарение остатка, дефлокуляцию с использованием KOH, центрифугирование, фильтрацию через сита 40 мкм, 10 мкм и 2 мкм и помещение образцов пыльцы в глицерин. Идентификация пыльцы и спор под световым микроскопом при увеличении 400х с использованием авторской справочной коллекции лаборатории геоэкологии Севера МГУ им. М. В. Ломоносова, российской базы данных пыльцы (pollendata.org.) и определителей.
Проведенный авторами анализ результатов исследований различных специалистов показал отсутствие единых позиций для адекватной оценки особенностей летнего температурного режима в Северной Якутии в позднем плейстоцене. Полученные в статье данные позволяют уточнить палеотемпературные реконструкции.
Палинологическая характеристика отложений разреза Зеленый Мыс фиксирует примерно семь среднепериодных циклических изменений растительного покрова, как в локальном, так и в региональном масштабе. Это выразилось в резкой смене доминантов: палиноспектры с преобладанием пыльцы карликовой березки с участием пыльцы сосны обыкновенной, сменяются палиноспектрами с доминированием пыльцы злаков и полыни или с доминированием недоразвитой пыльцы разнотравья и спор плаунка сибирского. При этом в пределах выделенных палинозон преимущественно сохраняется характер циркуляции атмосферы в весенне-летний период, циклы отражают изменения базиса эрозии и колебания температур вегетационного периода, а также его длительность.
Проведённая палинологическая характеристика разреза едомных отложений Зеленый Мыс позволила авторам выделить на диаграмме трех локальных палинозон: Varia + Artemisia (V+A), (интервал 27.0–24 м) – Varia + Selaginella sibirica (V+S) (интервал 24–18 м) и Pinus + Betula sect. Nanae (P+B), (интервал18.0–9.5 м). Практически все палиноспектры отражают открытые ландшафты мозаичных гипоарктических тундр. Реконструкция сумм положительных температур показала, что палиноспектры отражают троекратные колебания условий вегетации: два кратковременных периода существенного ухудшения условий вегетации (сумма положительных температур около 200-300 град. × дней), около 46, 34-33 тыс. кал. лет. (и два периода улучшения условий вегетации 45–43 тыс. кал. лет (сумма положительных температур около 600 град. × дней) и 32 тыс. кал лет (сумма положительных температур около 750 град. × дней).
Стиль изложения материала соответствует принятому уровню для научных статей. Принятая структура позволяет последовательно и аргументировано изложить методы и результаты исследований. Материал изложен хорошим научным языком, понятным для специалиста языком. Рубрикация текста адекватна содержанию и соответствует требованиям журнала.
Библиография включает 22 источников, что позволяет рассмотреть аргументацию различных авторов по рассматриваемой в статье теме.
В статье приведён подробный анализ позиций различных авторов по теме особенностей летнего температурного режима в Северной Якутии в позднем плейстоцене. Тон дискуссии с оппонентами уважительный, аргументация убедительна.
Рассматриваемая в статье тема представляет интерес для палеогеографов, палеоботаников, геокриологов, климатологов, а также для широкого круга читателей, интересующихся северной проблематикой. Работа имеет научную и практическую ценность, поэтому может быть опубликована в журнале «Арктика и Антарктика».
|
 Рус
Рус


















