|
DOI: 10.7256/2453-8922.2017.1.22323
Дата направления статьи в редакцию:
16-03-2017
Дата публикации:
02-04-2017
Аннотация:
Голоценовые повторно-жильные льды широко распространены в поймах рек озер и на лайдах, а также в торфяниках Ямала. Предметом исследования данной статьи явилась палинологическая характеристика разрезов голоценовых полигонально-жильных комплексов с повторно-жильными льдами в пределах различных геоморфологических уровней. Оценивалась надежность радиоуглеродного датирования полигонально-жильных комплексов на основании палинологической характеристики, а также рассмотрена возможность оценки сингенетичности повторно-жильных льдов на основании сопоставления палиноспектров льдов и вмещающих их отложений. Проведено выделение этапов развития растительного покрова в данном районе. Базовый подход к изучению полигонально-жильных комплексов -изучение палиноспектров повторно-жильных льдов и вмещающих их отложений, основан на том, что палиноспектры льдов и отложений отражают разные сезоны аккумуляции пыльцы. Если на поверхности субфоссильный палиноспектр формируется в течение всего сезона вегетации и отражает как региональные, так и локальные особенности растительного покрова, то палиноспектры повторно-жильных льдов в основном формируются из весеннего пыльцевого дождя, отражая в основном региональные характеристики. Оценка надежности полученных радиоуглеродных датировок базируется на оценке содержания переотложенных доплейстоценовых пыльцы и спор в датированном образце, что косвенно отражает содержание древней переотложенной органики. Сопоставление палиноспектров вмещающих отложений и повторно-жильных льдов также дает возможность оценить сингенетичность формирования повторно-жильных льдов. Выделены основные этапы развития растительного покрова на полуострове Ямал в голоцене. В изученных голоценовых разрезах Ямала с синкриогенными повторно-жильными льдами зафиксированы среднепериодные контрастные изменения структуры спорово-пыльцевых спектров, выразившиеся в резкой смене палинодоминантов: палиноспектры с преобладанием пыльцы осок сменяются палиноспектры с преобладанием пыльцы злаков, с преобладанием полыни – доминированием маревых, с ольховником и карликовой березкой – палиноспектрами с пыльцой древовидной березы, с кедровый стланником – палиноспектрами с кедром сибирским, с большим содержанием недоразвитой пыльцы двудольных трав – спектрами с доминированием зеленых мхов. Основная периодичность смены палиноспектров соответствует временным интерваламдлительностью 1,5-2 тыс. лет. Как правило, это связано с резким изменением условий вегетации, обусловленных изменениями базиса эрозии, облачности,направления и скорости ветра, влажности воздуха, различиямив мощности снежного покрова, температурных условий и длительности вегетациононного периода.
Ключевые слова:
голоцен, палиноспектры, повторно-жильный лед, радиоуглеродный возраст, полуостров Ямал, торфяник, палинологическая фаза, спорово-пыльцевая диаграмма, этапы развития растительноcти, климатические колебания
УДК: 551.89
Работа выполнена при финансовой поддержке РФФИ - грант № 17-05-00794 – А.
Abstract: Holocene wedge ices are widespread within laidas, peat bogs and flood plains of lakes and rivers of the Yamal Peninsula. The aim of the paper is to study palynological characteristic of exposures of wedge ices located at different geomorphological levels. The author estimates realibility of radiocarbon dating of polygonal wedge ices of the base of palynological characteristic and considers the possibility of estimating the idiogenousness of wedge ices on the basis of comparison of palinospectra of wedge ices and their deposits. The author detects several stages of vegetation evolution in this region. The main approach to ice wedge complex study is based on the fact that palynospectra of ices and deposits represent different seasons of pollen accumulation. While subfossil palynospectrum is accumulated during the whole vegetation season and consisits both of local and regional peculiarities of land cover, palinospectra of wedge ices are formed of spring pollen rains and reflect mainly regional characteristics. Reliability of the radiocarbon dating is estimated according to the constitution of Pre-Quaternary pollen in the sample, because it indicates the constitution of aicient penecontemporaneous organic material in the sample indirectly. Main stages of vegetation cover development in Holocene are distinguished on the base of these studies.
Keywords: Holocene, palynospectra, wedge ice, radiocarbon age, the Yamal Peninsula, peat, palynology phase, sporo-pollen plot, stages of vegetation development, climatic fluctuations
ВВЕДЕНИЕ
Голоценовые повторно-жильные льды широко распространены в поймах рек озер и на лайдах, а также в торфяниках Ямала (рис. 1). Однако их возраст и условия формирования не всегда определяются однозначно из-за высокой концентрации более древнего, в том числе и палинологического, органического материала. Для оценки зависимости содержания переотложенных форм в палиноспектрах повторно-жильных льдов и вмещающих отложений и датировок торфа автором был исследован ряд голоценовых торфяников Ямала, где переотложение доплейстоценовых палиноморф определяется достаточно надежно.
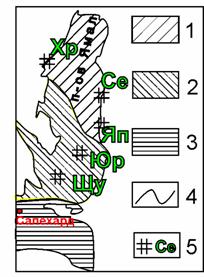
Рис. 1. Схема расположения на Ямале полигональных торфяников изученных автором:
Геокриологические зоны: 1 – 2 – зона сплошного распространения многолетнемёрзлых пород: 1 – среднегодовая температура пород ниже –5 °С, 2 – температура от –3 до –5 °С, 3 – зона массивно-островного и островного с поверхности распространения многолетнемёрзлых пород, среднегодовая температура пород от 0 до –3 °С; 4 – границы мерзлотных зон; 5 – расположение полигональных торфяников: Щу – Щучьинского, Яп – Яптиксалинского, Юр – Юрибейского, Се – Сеяхинского, Хр – Харасавейского
Автор предположила, что если в процессе палинологического анализа выявляются переотложенные формы можно полагать, что и остальная органика будет в заметной степени переотложенной и, следовательно, 14С датировка образца в этих случаях будет фиктивно более древней. Автором лично отобраны образцы и исследованы пыльца и споры в повторно-жильных льдах и вмещающих их торфяных отложениях голоценовых полигонально-жильного комплексов: Щучьинского, Яптиксалинского, Юрибейского, Сеяхинского (в устье р.Сеяха, Зеленая) и Харасавейского (см. рис. 1). Мощные полигональные торфяники на Яммале встречаются либо на поймах и лайдах, либо в озерно-болотных вкладках на более высоких элементах рельефа. Они накапливаются с разной скоростью которая определяетя как условиями накопления собственно торфа, так и льдистостью торфа. При высокой льдистости и благоприятных условиях дляроста и отмирания растений торфообразователей скорость их накопления может превышать несколько миллиметров в год.
Палинологическая характеристика и хронология голоценового Щучьинского полигонально-жильного комплекса
Мощный торфяник (5 - 5,5 м) расположен в долине р.Щучья на Южном Ямале (67°10’ с.ш., 69°5’ в.д.). Палинологически разрез изучался несколько раз. Л.В.Голубева [1] опубликовала результаты исследования в 1960 г. Этот разрез автором исследовался дважды (рис. 2, 3) с интервалом в 15 лет.

Рис. 2. Мощный полигональный торфяник в долине р.Щучья, Южный Ямал. Фото Ю.Васильчука

Рис. 3. Верхняя часть мощного полигонального торфяника в долине р.Щучья (а) и повторно-жильный лед (б) в его основании. Фото Ю.Васильчука (а) и А.Васильчук (б)
Самым выразительным геокриологическим элементом разреза является обнажение крупных (высотой более 5 м) сингенетических повторно-жильных льдов, рассекающих всю толщу торфа (рис. 4а) и проникающих в подстилающие торф озерные супеси и глины (рис. 4б).
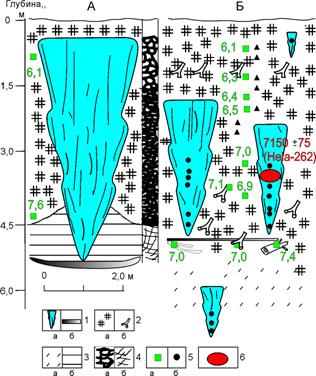
Рис. 4. Мощные ледяные жилы в голоценовом торфянике в долине р. Щучья: А) крупные повторно-жильные льды в центральной части разреза, изученные в 1977 г.; Б) небольшие погребенные повторно-жильные льды в краевой части торфяника, изученные и опробованные в 1997 г. Условные обозначения: 1 – лед: (а) повторно-жильный, (б) сегрегационный; 2 – растительные остатки: (а) торф, (б) стволы, ветки и корни берез; 3 – (а) супесь, (б) глина; 4 – криогенная текстура: (а) базальная, (б) тонкошлировая сетчатая; 5 – места отбора образцов: (а) органики на радиоуглеродный анализ, (б) повторно-жильных льдов на спорово-пыльцевой анализ; 6 – отбор стебелька мха для AMS-датирования и радиоуглеродная дата
В основании торфяника залегает мощный (до 1,5 м) древесный горизонт, сложенный, главным образом, стволами березы диаметром 0,2–0,3 м. Встречены стволы лиственницы, а также шишки ели и лиственницы. Присутствие корней и прикорневых веток свидетельствует об автохтонном характере залежи.
Разрез сингенетических повторно-жильных льдов в низовьях р.Щучья имеет принципиально важное значение для восстановления истории палеогеографического и геокриологического развития не только Ямала, но и Западной Сибири в целом. Его весьма южное положение (достаточно близкое к полярному кругу) позволяет установить практически самый южный предел развития сингенетических повторно-жильных льдов в эпоху голоценового оптимума.
Детальные радиоуглеродные определения позволили датировать возраст торфяника периодом голоценового «оптимума» (рис. 5). Период оптимума голоцена для Ямала установлен, как время иммиграции в тундру древесной растительности от 9,2 до 4,3 тыс. лет назад [2–6].
Примечательно то, что первые две радиоуглеродные датировки были получены около 40 лет назад в радиоуглеродной лаборатории Ленинградского университета. Тогда отбор образцов был выполнен из центральной части торфяника сотрудниками НПО «Аэрогеология». Две датировки 7680 ± 110 лет (ЛУ-1081) у основания и 6140 ± 80 лет (ЛУ-1082) в верхней части, были получены из центральной части торфяника. Радиоуглеродная AMS-датировка стебелька мха, взятого из осевой части ледяной жилы – 7150 ± 75 лет (Hela – 262), продемонстрировала, что формирование льда началось сразу же после начала накопления торфа.
Эта AMS датировка прямо подтвердила, что жилы активно росли в оптимум голоцена [6]. Параллельный анализ спорово-пыльцевых остатков в ледяных жилах и во вмещающих их отложениях выявил сходство в распределении содержания пыльцы деревьев.
Датировка 6110 ± 110 лет (Hel-4138) получена на глубине 0,7-0,8 м по образцу 364-YuV/67. Пыльца деревьев составляет 27%, встречена пыльца лиственницы (4%), кедра (4%), сосны (3%), ели (1%), ольхи (2%), березы (10%). Пыльца кустарников составляет 18%, это пыльца карликовой березки (12%), ольховника (4%), ивы (1%). Пыльца трав составляет 28%, пыльца злаков и осок содержится в равном соотношении - 9%, пыльца верескоцветных составляет 2%, полыни 1%, разнотравья 5%. Среди спор (25%) преобладают споры зеленых мхов (15%), участвуют споры сфагновых мхов (4%), а также споры Huperziaselago, Equisetum. Эта датировка выполнена по автохтонному материалу и близка к истинной.
Образец 364-YuV/74 отобран на глубине 1,35-1,4 м и датирован 6300 ± 100 лет (Hel-4139). Переотложенная пыльца содержатся в количестве 2%, хорошо заметна по степени метаморфизма и представлена пыльцой кувшинки, ореховых, тсуги.
Пыльца деревьев составляет 36%. Это пыльца лиственницы (2%), кедра сибирского (2%), сосны обыкновенной (1%), ели (2%), ольхи (3%), березы (26%). Пыльца кустарников составляет 15%, доминирует пыльца карликовой березки (11%), подчиненное значение имеет пыльца ольховника (3%) и ивы (1%). В состав пыльцы разнотравья входит пыльца вахты трехлистной, гречишных, гвоздичных, подорожниковых, зонтичных и др. Среди спор (19%) доминируют споры зеленых мхов (13%), споры сфагновых мхов составили 3%, встречены споры многоножковых папоротников и хвощей. Здесь, скорее всего также датирован автохтонный материал и датировка валидна.
Образец 364-YuV/48 отобран на глубине 2,7-2,8 м и датирован 7020 ± 100 лет (Hel-4135). Переотложенные пыльца и споры составили здесь менее 1%. При исследовании под микроскопом обнаружено значительное количество углистых частиц (возможно результат пожара), и чешуйки рыб. Среди пыльцы деревьев (51%) встречено много пыльцы лиственницы 7%, пыльца ели составила 5%, ольхи 12%, абсолютно преобладает пыльца березы (28%). Пыльца кустарников (8%) представлена пыльцой карликовой березки (6%), ольховника (1%) , ивы (1%). Пыльца трав и кустарничков (15,6%) представлена злаками (7%), осоками (9%), верескоцветными (2%), разнообразен состав пыльцы разнотравья (5%): бобовые, лютиковые, первоцветные, лилиецветные, яснотковые, вахтовые. Среди спор (15%) преобладают сфагновые мхи (8%), зеленые мхи составляют 4%, хвощи 3%. Эта датировка видимо несколько удревнена за счет водного привноса органики непыльцевой размерности.
Образец 364-YuV/45 отобран на глубине 3,6-3,7 м и датирован 6960 ± 100 лет (Hel- 4135). Переотложенные пыльца и споры здесь не встречены, отмечены остатки клещей-орибатид и насекомых.
Пыльца древесных (60%) представлена в основном пыльцой нескольких видов берез (27%) и ольхи (29%), пыльца хвойных единична: ель менее 1%, кедр сибирский 3%. Пыльца кустарников (20%), представлена ольховником (14%), карликовой березкой (6%), ивой – менее одного процента. Состав пыльцы трав очень беден, это только пыльца злаков (3%), верескоцветных (2%), полыни (2%) и маревых (1%). Состав спор (8%) наоборот разнообразен. Большую часть составляют споры хвощей (4%), отмечены также споры сфагновых мхов, зеленых мхов, разных видов плаунов и папоротников. Эта датировка вполне валидна.
В этом разрезе для нас интересна небольшая инверсия двух датировок, полученная в основании разреза. Наличие чешуек рыб свидетельствует о поступлении части органического материала из реки, что могло повлиять на верхнюю радиоуглеродную датировку в результате частичного привноса аллохтонного материала. Как видно из ряда датировок, даже небольшая примесь древней несинхронной органики оказывает воздействие на результаты датирования, и приводит к небольшим инверсиям в распределении датировок по разрезу.
В самой верхней части торфа на глубинах 0,7-0,8 м встречено сравнительно высокое (до 6% от общего количества) содержание переотложенных пыльцы и спор палеогенового возраста и высокое содержание деформированной пыльцы, что указывает на привнос переотложенных пыльцевых зерен вместе с минеральной составляющей.
Одним из индикационных признаков, согласно которому ряд исследователей предполагал деградацию мерзлых толщ на юге Ямала [7] было наличие пыльцы и макроостатков вахты трехлистной. Динамика популяции вахты трехлистной (Menyanthes trifoliata L.) отражает условия обводнения Щучьинского торфяника. Максимум содержания ее пыльцы отмечен на глубине 1,5–1,8 м. В момент формирования этого горизонта торфа, вахта трехлистная была распространена на значительной площади. Несколько ниже или выше по разрезу пыльцевые зерна вахты встречаются лишь в единичных количествах. По всей видимости, популяция вахты на поверхности торфяника сохранялась почти все время его формирования, что определялось повышенной увлажненностью его поверхности. Вахта исчезла на периферийных участках торфяника недавно, ее нет на глубине до 0,3–0,4 м и выше. Отметим, что и на поверхности современного осушенного торфяника она не встречена, зато вдоль берегов почти всех зарастающих водоемов и в том числе вдоль берегов мелких озер на пойме р.Щучья вахта трехлистная отмечается в виде узкой полосы. В поверхностных современных пробах пыльца вахты трехлистной отмечена в единичных количествах, что отражает преимущественно субаэральный режим поверхности современного торфяника.
А.П.Тыртиков [8, 9] отмечал многочисленные находки вахты даже в районе пос. Гыда, т.е. в районе с очень низкой температурой многолетнемерзлых толщ (до –10 °С). Действительно массовая встречаемость вахты трехлистной действительно ограничивается на Ямале южными районами [10], обычно располагается южнее 68° с.ш., однако отдельные фрагменты ее ареала могут встречаться значительно севернее.
На севере Европейской части России вахта трехлистная обитает на всем пространстве тундровой зоны вплоть до побережья Арктики, а в Средней и Восточной Сибири она иногда обильно встречается севернее 70° и даже севернее 72° с.ш. [10]. Это вполне связано с тем, что северная граница ее ареала регламентируется летними, а не среднегодовыми температурами). В разрезе торфяника на р.Щучья пыльца вахты прослежена на протяжении всего периода формирования торфяника, она, очевидно, фиксирует степень увлажнения массива и не может служить индикатором регионального протаивания. В.Я. Кац впоследствии [11] также признавал, что обнаруженные им в низовьях Оби кора, древесина, пни ели, березы, лиственницы и остатки бореальных растений указывают на то, что именно летние (а не среднегодовые) температуры были выше современных на 3,5–4 °С.
На возможно значительно более южное положение границы протаивания в оптимум голоцена, как упомянуто выше, указывали и А.И.Попов [12] и И.Д.Данилов [13]. Полученные Ю.К.Васильчуком [5, 6] изотопные данные по синкриогенным повторно-жильным льдам указывают, что здесь этот этап следует трактовать как время повышенной континентальности климата: лето несколько теплее современного (в среднем на 1–3 °С), а зима в среднем на 2–3 °С холоднее.
Это и могло вызвать увеличение интенсивности морозобойного растрескивания и связанного с ним повторно-жильного льдообразования, как в органогенных, так и в минеральных аллювиальных и аллювиально-морских толщах. Более 10 радиоуглеродных датировок, выполненных по вмещающему мощные сингенетические жилы торфу, демонстрируют очень быстрый рост торфяника (около 5 м за период менее тысячи лет).
Итак, оптимум – это период с относительно теплым летом, продвижением вплоть до 71-72° с.ш. лиственницы и березы, смещение на север границ прерывистой мерзлоты и одновременное формирование сингенетических жил в непосредственной близости от зоны прерывистой мерзлоты.
Л.В.Голубевой [1] была получена детальная палинологическая диаграмма по Щучьинскому торфянику (рис. 5).
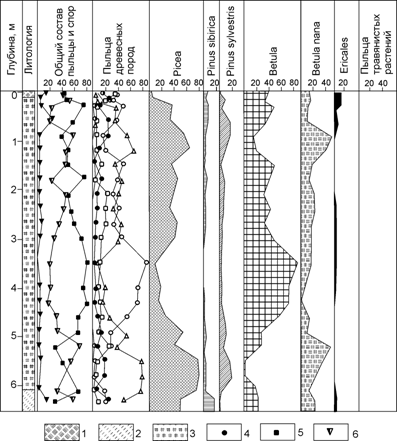
Рис. 5. Спорово-пыльцевая диаграмма разреза Щучьинского голоценового торфяника (по Л.В.Голубевой [1]), 1 – талый торф; 2 – супесь; 3 – мерзлый торф; 4 – пыльца трав и кустарничков; 5 – пыльца деревьев и кустарников, 6 – споры
Выраженность хода кривых содержания пыльцы древесных пород позволили выделить, нижний максимум ели, максимум пыльцы кустарниковых берез, второй максимум ели. Это стало основанием для вывода о том, что торфяник накапливался в течение всего голоцена, по крайней мере, последние 6-8 тыс. лет.
Хотя прослеженная Л.В.Голубевой смена доминирования отдельных таксонов близка к стандартной для Западной Сибири, она имеет, согласно полученным нами [3, 5, 6] радиоуглеродным датировкам, иной временной масштаб. Г.М.Левковская [14] отмечала, что палинологическая характеристика Щучьинского торфяника несколько отличается от остальных диаграмм Западной Сибири, некоторые стадии здесь не отмечаются. После проведения подробного радиоуглеродного датирования стало очевидно, что этот торфяник накопился в десять раз быстрее, т.е. за 700-1000 лет.
Сопоставление диаграммы по вмещающим отложениям на р.Щучья, составленной автором (рис. 6, табл. 1), и диаграммы, полученной по повторно-жильным льдам (рис. 7, табл. 2), выявило, что наблюдается существенное сходство с диаграммой Л.В.Голубевой (сравн. рис. 5 и 6).
Общими моментами являются нижний максимум пыльцы древовидной березы, верхний максимум пыльцы кедра сибирского и сосны обыкновенной, совпадающий с максимумом пыльцы верескоцветных. Особенно наглядно это сходство проявилось в диаграмме из повторно-жильных льдов: доминирование пыльцы ели в составе регионального пыльцевого дождя и выраженный максимум пыльцы верескоцветных. Таким образом, региональный пыльцевой дождь действительно отразил динамику смены растительных формаций.
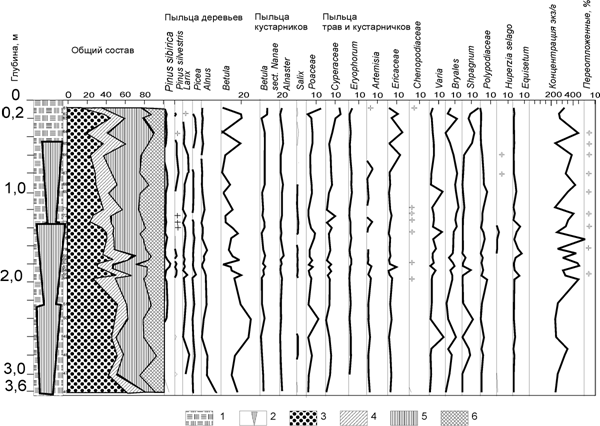
Рис. 6. Спорово-пыльцевая диаграмма Щучьинского торфяника (сборы 1977 г., палинологический анализ Е.А.Петровой и А.К.Васильчук): 1 – торф; 2 – повторно-жильный лед; 3 – пыльца древесных; 4 – пыльца кустарников; 5– пыльца трав и кустарничков; 6 – споры
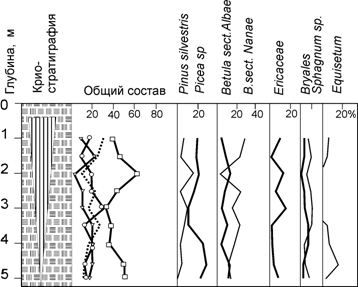
Рис. 7. Спорово-пыльцевая диаграмма из повторно-жильных льдов, сформировавшихся в толще торфяника р.Щучья на юге п-ова Ямал
Таблица 1. Спорово-пыльцевые спектры голоценового полигонально-жильного торфяника на р.Щучья, южный Ямал (вмещающие отложения, сборы 1977 г.)
|
Точка 129–YuV
|
0
|
1
|
2
|
4
|
5
|
6
|
7
|
9
|
10
|
11
|
11а
|
12
|
|
Глубина отбора, м
|
0,2
|
0,5
|
1,0
|
1,5
|
1,6
|
1,7
|
1.8
|
2,6
|
2,9
|
3,1
|
3,2
|
4,0
|
|
Пыльца деревьев
|
24
|
47
|
44
|
32
|
20
|
30
|
26
|
31
|
32
|
25
|
22
|
23
|
|
Пыльца кустарников
|
30
|
28
|
8
|
21
|
17
|
9
|
29
|
12
|
5
|
17
|
4
|
1
|
|
Пыльца трав и кус–ничк.
|
16
|
16
|
26
|
10
|
31
|
31
|
17
|
19
|
55
|
29
|
47
|
22
|
|
Споры
|
30
|
9
|
22
|
37
|
32
|
30
|
28
|
38
|
8
|
29
|
27
|
54
|
|
Picea sp.
|
4
|
7
|
14
|
11
|
10
|
12
|
6
|
18
|
6
|
6
|
7
|
8
|
|
Pinus sibirica
|
6
|
7
|
3
|
3
|
2
|
1
|
2
|
1
|
–
|
4
|
1
|
2
|
|
Pinus silvestris
|
3
|
1
|
4
|
1
|
2
|
2
|
–
|
–
|
–
|
3
|
–
|
1
|
|
Alnus sp
|
–
|
6
|
3
|
3
|
1
|
3
|
2
|
5
|
1
|
3
|
1
|
2
|
|
Betula sect. Albae
|
11
|
26
|
20
|
14
|
5
|
12
|
16
|
7
|
25
|
9
|
13
|
10
|
|
Betula sect. Nanae
|
27
|
26
|
7
|
19
|
16
|
8
|
29
|
12
|
4
|
16
|
3
|
1
|
|
Salix sp
|
2
|
–
|
1
|
1
|
1
|
1
|
–
|
–
|
1
|
–
|
–
|
–
|
|
Alnaster sp.
|
1
|
2
|
–
|
1
|
–
|
–
|
–
|
–
|
–
|
1
|
1
|
–
|
|
Poaceae
|
1
|
6
|
1
|
1
|
1
|
29
|
5
|
6
|
5
|
3
|
3
|
1
|
|
Cyperaceae
|
5
|
5
|
20
|
6
|
26
|
–
|
5
|
6
|
5
|
3
|
3
|
1
|
|
Ericaceae
|
8
|
4
|
2
|
1
|
1
|
–
|
5
|
–
|
1
|
4
|
–
|
–
|
|
Varia
|
1
|
–
|
2
|
–
|
2
|
1
|
2
|
7
|
2
|
2
|
1
|
1
|
|
Artemisia sp.
|
1
|
1
|
1
|
1
|
1
|
1
|
1
|
2
|
1
|
1
|
–
|
1
|
|
Chenopodia-ceae
|
–
|
–
|
–
|
1
|
–
|
–
|
–
|
1
|
1
|
–
|
–
|
–
|
|
Bryales
|
4
|
4
|
3
|
5
|
3
|
5
|
5
|
9
|
2
|
16
|
2
|
3
|
|
Sphagnum sp.
|
24
|
4
|
13
|
21
|
11
|
10
|
9
|
1
|
2
|
16
|
2
|
3
|
|
Polypodia-ceae
|
–
|
–
|
–
|
1
|
–
|
–
|
–
|
–
|
–
|
1
|
–
|
1
|
|
Lycopodium sp.
|
–
|
–
|
–
|
–
|
–
|
–
|
–
|
3
|
–
|
1
|
–
|
1
|
|
Количество зерен, экз.
|
466
|
293
|
395
|
309
|
240
|
280
|
185
|
157
|
213
|
276
|
236
|
178
|
Таблица 2. Спорово–пыльцевые спектры голоценового повторно–жильного льда из полигонально-жильного торфяника на р.Щучья, южный Ямал (сборы 1977 г.)
|
Точка 128–YuV
|
11
|
10
|
12
|
13
|
14
|
15
|
16
|
17
|
18
|
|
Глубина отбора, м
|
1,0
|
1,5
|
2,0
|
2,5
|
3,0
|
3,5
|
4,0
|
4,5
|
5,0
|
|
Пыльца деревьев
|
40
|
45
|
66
|
45
|
36
|
39
|
38
|
50
|
49
|
|
Пыльца кустарников
|
28
|
23
|
10
|
23
|
19
|
27
|
19
|
14
|
17
|
|
Пыльца трав и кус–ничк.
|
18
|
10
|
19
|
19
|
32
|
14
|
22
|
15
|
17
|
|
Споры
|
14
|
22
|
5
|
13
|
13
|
20
|
21
|
21
|
17
|
|
Picea sp.
|
21
|
20
|
23
|
18
|
10
|
13
|
26
|
27
|
23
|
|
Pinus sibirica
|
7
|
3
|
18
|
10
|
12
|
9
|
4
|
6
|
–
|
|
Pinus silvestris
|
3
|
3
|
7
|
2
|
3
|
1
|
2
|
–
|
9
|
|
Alnus sp
|
3
|
2
|
–
|
3
|
4
|
1
|
–
|
–
|
–
|
|
Betula sect. Albae
|
6
|
16
|
18
|
12
|
7
|
15
|
6
|
17
|
17
|
|
Betula sect. Nanae
|
27
|
22
|
5
|
23
|
19
|
24
|
19
|
13
|
17
|
|
Salix sp
|
1
|
1
|
–
|
–
|
–
|
1
|
–
|
1
|
–
|
|
Alnaster sp.
|
–
|
–
|
5
|
–
|
–
|
2
|
–
|
–
|
–
|
|
Poaceae
|
4
|
4
|
6
|
10
|
12
|
4
|
4
|
4
|
6
|
|
Cyperaceae
|
1
|
–
|
–
|
–
|
–
|
1
|
3
|
1
|
–
|
|
Ericaceae
|
10
|
1
|
13
|
7
|
16
|
2
|
3
|
5
|
9
|
|
Varia
|
3
|
4
|
–
|
1
|
2
|
2
|
10
|
4
|
1
|
|
Artemisia sp.
|
–
|
1
|
–
|
1
|
2
|
5
|
–
|
–
|
1
|
|
Chenopodiaceae
|
–
|
–
|
–
|
–
|
–
|
–
|
2
|
1
|
–
|
|
Bryales
|
4
|
6
|
–
|
4
|
4
|
9
|
9
|
2
|
3
|
|
Sphagnum sp.
|
6
|
12
|
5
|
9
|
9
|
9
|
6
|
3
|
7
|
|
Polypodiaceae
|
–
|
1
|
–
|
–
|
–
|
1
|
–
|
–
|
2
|
|
Lycopodium sp.
|
–
|
–
|
–
|
–
|
–
|
–
|
–
|
–
|
–
|
|
Количество зерен, экз.
|
113
|
77
|
39
|
163
|
69
|
86
|
68
|
105
|
84
|
Выделенные палинологические фазы, длились сотни лет (так как возраст этого мощного торфяника не превышает 1 тыс. лет). Очевидно, что развитие и смена растительных ассоциаций подчиняются одним и тем же закономерностям, именно поэтому для палинологических построений так важна информация по радиоуглеродному датированию. Только на основании ряда датировок можно определить скорость накопления отложений и соответственно длительность палинофаз.
Отметим, что формирование повторно-жильных льдов в период 7200-6100 лет назад в районе р.Щучья маркируется по палинологическим данным повышением роли пыльцы ели, березы и верескоцветных в региональном пыльцевом дожде, в локальных фитоценозах на это время приходится возрастание содержания пыльцы высокоствольной березы, лиственницы и ели.
В средней части разреза отмечается повышение содержания пыльцы полыни и маревых (см. рис. 6). В целом отмечается последовательный рост полигонального торфяника в условиях развития березово-еловых лесов и завершение его развития из-за снижения грунтового питания, о чем свидетельствует исчезновение пыльцы вахты трехлистной и возрастание роли пыльцы верескоцветных, в частности подбела, спор сфагновых мхов.
Палинологическая характеристика и хронология голоценового Яптиксалинского полигонально-жильного комплекса
Севернее автором изучен торфяник на первой лагунно-морской террасе у пос. Яптиксале (рис. 8).

Рис. 8. Полигонально-жильный торфяник близ пос. Яптиксале (п-ов Ямал). Фото Ю.Васильчука
Содержащий древесину слой торфа в торфянике близ пос. Яптиксале датирован 8950 14С лет назад и древесина 8700 14С лет назад.
Торф в верхней части разреза датирован 1590 14С лет назад. Разрез можно визуально разделить на три части. Нижний слой торфа с древесными остатками в интервале 2-5,5 м, слой опесчаненного торфа со слабо выраженной горизонтальной слоистостью и верхний слой песка.
Если рассматривать региональные компоненты, то можно отметить существенное сходство палиноспектров в торфянике и в ледяных жилах близ пос. Яптиксале из интервала 2-4 м и палиноспектров торфяника на р.Щучьей из вмещающих отложений, так и из повторно-жильных льдов.
Однако имеются отличия, объяснимые в рамках представлений о формировании регионального пыльцевого дождя в тундровой зоне [15]. Здесь и во вмещающих отложениях и в повторно-жильных льдах мало пыльцы ели. Пыльца ели в заметных количествах обнаружена только в основании разреза.
В повторно-жильном льду доминирует пыльца березы древовидной и сосны обыкновенной (Betula sect. Albae 28%, Pinus silvestris 9%), отмечена, березки карликовой, полыни, разнотравья и спор сфагновых мхов.
Судя по палиноспектрам из повторно-жильного льда, ледяные жилки, обнаруженные в данном торфянике сформировались раньше, чем повторно-жильные льды в Щучьинском торфянике.
Палинологические данные по повторно-жильным льдам на первой лагунно-морской террасе у пос.Яптиксале (датируемые 8-7 тыс.лет назад) можно сопоставить с данными по голоценовому торфянику в долине р.Щучья (7-6 тыс.лет назад) (сравн. рис. 9 и рис. 6).

Рис. 9. Спорово-пыльцевая диаграмма и 14С датировки торфяника на первой лагунно-морской террасе у пос. Яптиксале на Ямале 1 – торф; 2 – песок; 3 – древесные остатки; 4 - повторно-жильный лед; 5 – радиоуглеродная датировка; 6 – споры; 7 – пыльца древесных пород; 8 – пыльца кустарничков и трав; 9 – пыльца сосны; 10 – пыльца ели; 11 – пыльца кедра; 12 – пыльца ольхи; 13 – пыльца лиственницы; 14 – пыльца березы; 15 – пыльца маревых; 16 – пыльца полыни; 17 – пыльца верескоцветных; 18 – пыльца карликовой березки (Betula sect Nanae); 19 – пыльца осок; 20 – пыльца злаков; 21 – пыльца ольховника; 22 – споры папоротников; 23 – споры сфагновых мхов (Sphagnum); 24 – споры зеленых мхов (Bryales); 24 – споры хвощей; 26 – споры плаунов
Очевидно, формирование повторно-жильных льдов у пос.Яптиксале происходило, когда в региональном пыльцевом дожде преобладала пыльца березы древесных форм. Прослеживается характерное изменение регионального пыльцевого дождя, выделенное во льду разреза на Щучьей в нижней части жил в интервале глубин 2,2-4,5 м. Пик содержания пыльцы карликовой березки совпадает с максимумом содержания пыльцы верескоцветных примерно в том же интервале глубин.
Очевидно, что по содержанию пыльцы и спор в голоценовых повторно-жильных льдах можно проследить изменения регионального пыльцевого дождя по всей территории Ямало-Гыданской области.
Палинологическая характеристика и хронология голоценового полигонально-жильного комплекса в долине реки Юрибей
Улучшение условий вегетации в арктических и гипоарктических тундрах привело к увеличению пыльцевой продуктивности типично тундровых растений в сочетании с невысокой продуктивностью древесных растений, находящихся на северном пределе обитания. Такая ситуация и сейчас наблюдается на севере лесотундры, например в районе разреза Щучья. Эта ситуация сложилась около 7 тыс. лет назад в среднем течении р. Юрибей. На первой надпойменной террасе р. Юрибей на Ямале, в содержащем древесину слое торфа, доминирует пыльца карликовой березки (до 40%) (рис. 10).

Рис. 10. Спорово-пыльцевая диаграмма и 14С датировки торфяника на первой надпойменной террасе р.Юрибей на Ямале:
1 – торф; 2 – супесь; 3 – радиоуглеродная датировка; 4 – пыльца древесных пород; 5 – пыльца кустарничков и трав; 6 – споры; 7 – пыльца ольхи; 8 – пыльца березы; 9 – пыльца сосны; 10 – пыльца кедра; 11 – пыльца ели; 12 – пыльца злаков; 13 – пыльца карликовой березки (Betula sect Nanae); 14 – пыльца ольховника; 15 – пыльца осок; 16 – пыльца полыни; 17 – споры сфагновых мхов (Sphagnum); 18 – споры зеленых мхов (Bryales); 19 – споры плаунов
Близкие по характеру изменения регионального пыльцевого дождя прослеживаются и на близкой к стандартной пыльцевой диаграмме по торфянику на мысе Каргинском (рис. 11) на Енисее.
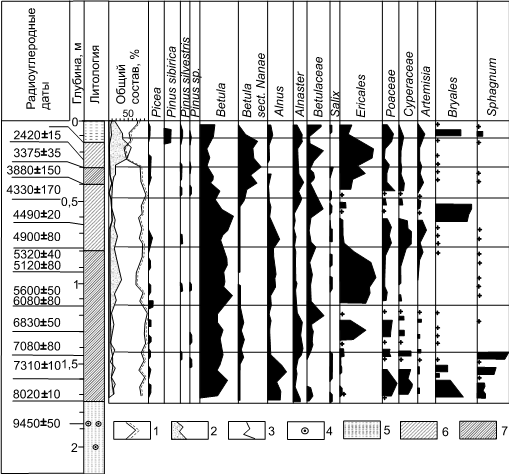
Рис. 11. Спорово-пыльцевая диаграмма и 14С датировки торфяника в верхней части 32-35-метровой террасы на правом берегу р. Енисей, 1,2 км выше мыса Каргинского (по [16]): а - основная часть спектра, б – дополнительная часть спектра: 1 – споры; 2 – пыльца трав и кустарничков; 3 – пыльца древесных пород; 4 – остатки бобровой плотины. 5 – 7 – торф. В разрезе получено 15 14С датировок: на глубине 0,13-0,23 м – 2420 ± 15 лет (СОАН-55); 0,23-0,30 м – 3375 ± 35 (СОАН-56); 0,3-0,4 м – 3880 ± 150 (СОАН-57); 0,4-0,5 м – 4330 ± 170 (СОАН-58); 0,5-0,65 – 4490 ± 20 (СОАН-63); 0,65-0,8 м – 4900 ± 80 (СОАН-64); 0,8-0,95 – 5320 ± 40 (СОАН-65) и 5120 ± 80 (СОАН-66); 0,95-1,17 – 5600 ± 50 (СОАН-67) и 6080 ± 80 (СОАН-68); 1,17-1,31 м – 6830 ± 50 (СОАН-59); 1,31-1,45 м – 7080 ± 80 (СОАН-60); 1,45-1,6 м – 7310 ± 10 (СОАН-61); 1,6-1,74 м – 8020 ± 10 (СОАН-62) и по бобровой плотине 9540 ± 50 лет (ГИН-260)
Она демонстрирует, что в период от 9,5-8 до 2,5 тыс. лет назад здесь происходили заметные ландшафтные изменения: около 3,6 тыс. лет назад появилась тундровая растительность, и березовые леса сменила лесотундра в первой половине суббореального периода [16]
Палинологическая характеристика и хронология голоценовых полигонально-жильных комплексов в устье реки Сеяха (Зеленая)
Автору удалось изучить палиноспектры повторно-жильных льдов и проследить палеогеографические особенности развития территории в голоцене на примере сингенетических повторно-жильных льдов в парагенезе с торфяными жилами, залегающими параллельно ледяным жилам (рис. 12) в голоценовой озерно-болотной вкладке на поверхности 24-метровой террасы р.Сеяха. В южной части обнажения сингенетические голоценовые повторно-жильные льды встречены в парагенезе с торфяными жилами. Хотя нельзя исключить, что это псевдоморфозы, так как помимо торфа в сложении грунтовых жил участвуют мелкие веточки и древесина березы с корой, для проникновения которых в первично грунтовые жилы требуется значительное раскрытие морозобойных трещин. Ледяные жилы залегают практически параллельно торфяным (рис. 13). Головы тех и других располагаются на одной глубине - около 0,5-0,7 м.
|
а
|
б
|
|

|

|
Рис. 12. Крупная ледяная жила в голоценовом торфянике в долинев устье р.Сеяха (Зеленая): до опробования (а) и после отбора образцов. Фото А.Васильчук
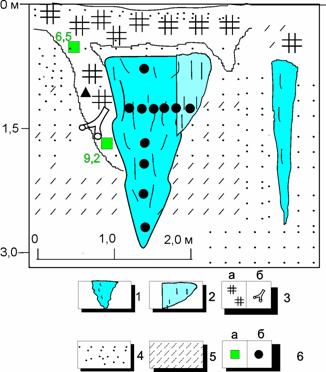
Рис. 13. Сингенетические повторно-жильные льды коричнево-розовые в парагенезе с торфяными жилами, в южной части торфяника, расположенного на поверхности третьей 24-метровой террасы в устье р.Сеяха (Зеленая): 1 – повторно-жильный лед; 2 – лед каймы; 3 – органические остатки в залежи: а – торф, б – ветки кустарников; 4 – песок; 5 – супесь, 6 – отбор образцов: а – на 14С, б – на спорово-пыльцевой анализ
Непосредственно над головой жил залегает серовато-коричневый оторфованный тонкослоистый песок (мощность прослоев 0,3-0,03 м), слоистость выражена колебаниями степени оторфования и изменением гранулометрического состава. Выше залегает слой коричневого мерзлого торфа, который похож на тот торф, который вместе с песком выполняет грунтовые жилы. Нижняя граница этого слоя неровная карманообразная (мощность прослоя 0,3 м). Ледяная жила, скорее всего, развивалась сингенетично накоплению грунтовой торфяной жилы. Об этом свидетельствует изгиб вверх слоев вмещающих слоистых песков на боковом контакте с ледяной жилой и спорово-пыльцевой спектр льда (см. рис. 13). Лёд жил розовато-коричневатый, ширина головы жилы более 1,6 м, на боковых контактах отмечается белый сахаристый лед (примерно 0,1 м в ширину). Торфяные жилы достигают 2 м по вертикали, они сложены мерзлым опесчаненным торфом слаборазложившимся с веточками с корой, листьями дриад, камнеломок и карликовой березки.
Вмещающие жилы отложения представлены желтоватым мелким оторфованным песком с включениями веточек и гнездами оторфования. Слоистость выражена вариациями цвета от темно-коричневого до желтовато-серого, а также степенью оторфования прослоев. Мощность этих прослоев от 0,15-0,01 м. Слои загибаются кверху и на контакте с ледяными, и с грунтовыми жилами. На это также указывает согласный характер залегания ледяной и торфяной жил. Образец торфа с маленькими веточками в торфяной жиле в верхней части разреза датирован 9280 ± 140 лет (Hel-4031). В центральной части массива также отмечен парагенез торфяных и ледяных жил. Здесь торфяные жилы, продолжают ледяные жилы снизу. Торфяная жила была датирована 9300 ± 100 лет (ГИН-2472). Таким образом, торфяные грунтовые жилы активно формировались в начальную фазу голоценового оптимума, определяемого для Ямала, как уже сказано, от 9,2 до 4,3 тыс. лет назад, в результате значительного иссушения слоя сезонного протаивания в начале голоценового оптимума.
Формирование повторно-жильных льдов по торфяным жилам в начале голоцена - явление характерное для северо-западного сектора Российской криолитозоны. Подобный парагенез описан нами в торфянике близ г. Воркута: 14С датировки древесины и торфа из вертикальной грунтово-торфяной жилы 9,2 тыс. лет [17]. На западе п-ва Ямал вблизи пос. Марресале торфяная жила на глубине 2,5 м опробована Ф.А.Каплянской и В.Д.Тарноградским и датирована [18] в 9010 ± 100 лет (ЛУ-1267). Во всех трёх случаях грунтовые торфяные жилы находятся в парагенезе с мощными ледяными жилами (высотой 3,5 м и 2 м), часто рассекая грунтовые жилы, т.е. повторно-жильный лёд формировался сразу после образования торфяной жилы и наследовал её. Суровые зимние условия были причиной промерзания накапливавшихся озерных и болотных отложений и роста повторно-жильных льдов. Условия вегетационного периода были напротив достаточно благоприятными для продвижения на север древесной растительности. Степень континентальности была настолько высока, что на 70о с.ш. появилась лиственница. Это подтверждается не только обнаружением пыльцы лиственницы (рис. 14) в верхней части жилы, залегающей на периферии и находкой веточек и сочлененных игл лиственницы в северном фрагменте обнажения на периферии мощного торфяника (рис. 15), остатков ее стволов не обнаружено.
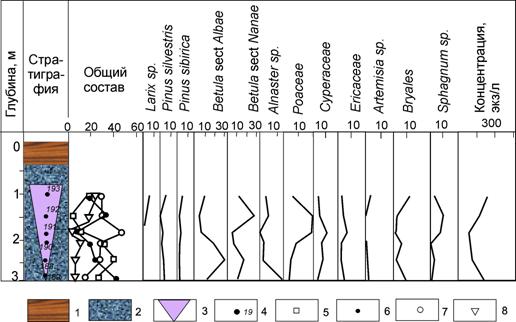
Рис. 14. Спорово-пыльцевая диаграмма голоценового коричневато-розового повторно-жильного льда в парагенезе с торфяными жилами, расположенного в южной части торфяника, на поверхности третьей 24-метровой террасы в устье р.Сеяха (Зеленая): 1 – торф; 2 – оторфованная супесь; 3 – повторно-жильный лед; 4 – точки отбора образцов; 5 – пыльца деревьев; 6 – пыльца кустарников; 7 – пыльца трав и кустарничков; 8 – споры;

Рис. 15. Полигональный голоценовый торфяник в устье р.Сеяха (Зеленая). Фото Ю.Васильчука
В самом нижнем слое черного обугленного торфа, вероятно связанного с пожаром, веточки ивы и березы с белой корой датированы в 9110 ± 120 лет (Hel-4057). Выше залегающий торф вокруг обугленных веток датируется 9020 ± 130 лет (Hel-4059), обугленные веточки из верхнего прослоя торфа имеют датировку 8940 ± 130 лет (Hel - 4058) и иглы лиственницы из гнезда торфа над верхним торфяным прослоем датированы на тандемном масс-спектрометре в 5990 ± 80 лет (Hela-200).
Палинологическая характеристика повторно-жильного льда коричнево-розового цвета, расположенного в первом - южном фрагменте торфяника даёт представление о весеннем пыльцевом дожде, формировавшемся здесь 6-9 тыс. лет назад. Содержание пыльцы древесных колеблется от 41 до 8% (см. рис. 14).
Пыльца сосен и ольхи представлена в небольших количествах и не превышает современные фоновые значения для данного района.
Главной особенностью полученных палиноспектров является высокое содержание пыльцы древесных видов берез 6-36% и встреченная в верхней части жилы пыльца лиственницы 1-5%. Пыльца ольховника доминирует в нижней части жилы. Среди пыльцы трав основную роль играет пыльца злаков (5-32%), встречается пыльца осок и пушицы. Довольно разнообразен для палиноспектров повторно-жильных льдов состав пыльцы разнотравья. В верхнем образце обнаружена пыльца ряски - обитателя мелких озер. Здесь встречена пыльца полыни и маревых. Палиноспектры отличаются от палиноспектров современной лесотундры высоким содержанием пыльцы злаков. Формирование повторно-жильных льдов пришлось на период, когда на широте пос. Сеяха в подходящих экотопах появились отдельные рощи из высокоствольной березы и, впоследствии, лиственницы. О том, что лиственница могла участвовать в фитоценозах, свидетельствует обнаружение пыльцы лиственницы в верхнем фрагменте жилы. Это подтверждается датировкой иголки лиственницы из гнезда торфа 5990 ± 80 лет (Hela-200). Следовательно, отдельные лиственницы могли встречаться на широте р.Сеяха (Зеленая) около 6 тыс. лет назад. Отсутствие стволов лиственницы может быть обусловлено пожарами, следы которых отмечены в разрезе, и тем, что лиственница на северном пределе своего обитания имеет рыхлую древесину, которая быстро разрушается. Обычно среди остатков встречаются веточки, иглы и шишки и отсутствует древесина. Мы полагаем, что повторно-жильные льды формировались в течение здесь 8,5-6 тыс. лет назад в течение одного цикла изменения направления и скорости ветра, когда доминирование пыльцы древовидной березы с участием пыльцы ольховника при относительно низкой концентрации пыльцы сменилось доминированием пыльцы злаков и, затем, пыльцы карликовой березки в весеннем пыльцевом дожде.
В палиноспектрах льда этих жил запечатлен рост сумм положительных температур в период от 8,3 до 6 тыс. лет.
Обобщение палинологических и изотопных данных позволило реконструировать ландшафтно-климатическую обстановку в этом регионе. Установлено три этапа развития растительности и климата: относительно теплый и влажный этап с современной тундровой растительностью, этап холодного и сухого климата, характеризуемый усилением роли ксерофильной растительности, наиболее теплый этап, относящийся к началу предбореального периода голоцена, когда кустарники продвинулись максимально на север до 74° с.ш.
Таким образом, раннепредбореальное потепление проявилось на арктическом побережье как термический оптимум в условиях еще континентального климата, хотя и значительно менее сурового, чем в позднеледниковое время. Все последующие голоценовые потепления происходили уже в условиях морского типа климата, благодаря сглаживающему влиянию холодных вод Северного Ледовитого океана. Даже максимальное потепление 4500 - 6000 лет назад, проявившееся как климатический оптимум во многих регионах Северного полушария [19], оказало меньшее воздействие на природу Арктики.
Раннеголоценовый климатический оптимум в высоких широтах Арктики отмечают многие исследователи [20, 21, 22]. Главной причиной того, что это потепление оказалось максимальным для современных приморских и островных районов Арктики, является быстрое повышение уровня Мирового океана к началу бореального периода и, как следствие, превращение районов с резко континентальным климатом в островные и приморские территории с соответствующими климатическими условиями.
Этапы развития растительного покрова, торфяников
и повторно-жильных комплексов Ямала в голоцене
Палиноспектры с такими характеристиками продуцируют биоценозы, функционирующие в течение короткого летнего периода, что характерно для позднего плейстоцена. Торф, залегающий в верхней части разреза, по нашему мнению является позднеплейстоценовым, а датировка, полученная по пузырькам метана из жильного льда, свидетельствует о проникновении голоценового льда в позднеплейстоценовый.
Можно говорить об общих и локальных особенностях развития растительности, торфяников и повторно-жильных льдов в голоцене. Общие черты обусловлены глобальным изменением теплообеспеченности, в частности заметным увеличением температур вегетационного периода, при сохраняющихся суровых зимних условиях. Локальные черты связаны с неравномерным увлажнением территории, особенно важном факторе для криолитозоны, так как с этим связан и неравномерный характер промерзания массивов и различная интенсивность льдонакопления. Повторно-жильные льды формировались в течение всего голоцена по всей территории Ямала. Естественно, что палиноспектры из отложений, вмещающих повторно-жильные льды, весьма разнообразны.
1 этап: 10 - 8,7 тыс. лет назад. Раннеголоценовое летнее потепление оказалось максимальным для северных районов Ямала из-за того, что к началу бореального периода произошло быстрое повышение уровня Мирового океана и, как следствие, на этих территориях климат из резкоконтинентального стал морским.
В криолитозоне этот этап практически фиксирует начало термического оптимума голоцена, с его началом связано увеличение летних температур и вызванное им повсеместное увеличение глубины сезонного протаивания на поверхности террас. Основной лесообразующей породой в северных форпостах леса на Ямале была береза. Низкие зимние температуры (возможно даже более холодные зимы, чем отмечаемые сейчас) приводили к интенсивному росту повторно-жильных льдов на поверхности торфяных болот. Иссушение верхних горизонтов в отдельные наиболее теплые летние сезоны вело к активизации пожаров. На спорово-пыльцевых диаграммах голоценовых полигонально-жильных комплексов Ямала этот этап может быть выделен по локальному максимуму пыльцы березы sect. Albae, участием пыльцы ели. В это время отмечается очень невысокое содержание пыльцы кустарниковых берез, ольховника и ивы при значительном количестве пыльцы полыни, маревых и гвоздичных.
2 этап: 8,7 - 7,8 тыс. лет назад. Начало этапа отмечено дренированием озерных котловин и активным заселением их древесной растительностью - это своеобразные мини-рефугиумы в которых деревья защищены от ветра. Это было благоприятнымдля развития полноценных стволов диаметром до 30-40 см. Возможно, что, в пределах выделяемого на изотопных диаграммах гренландского ледникового щита события 8к, условия как летнего, так и зимнего периода были близки к позднему плейстоцену: резкий переход к положительным температурам и практическое отсутствие осадков летом. Произрастание деревьев в период 8,4 - 8,2 тысяч лет назад в пределах озерно-болотных котловин вероятно, напоминало картину "пьяного" леса. Периодический подъем зеркала вод приводил к отмиранию древесной растительности и ее захоронению.
Деревья в этот период сохранялись лишь на кочках или на небольших мерзлых буграх в пределах котловин и по их окраинам. Рост деревьев на поверхности котловин и в непосредственной близости от них около 8 тысяч лет назад прекратился. Заполнение котловин мхами, осоками и остатками трав и кустарничков происходило очень быстро, не только за счет активного роста болотной растительности "in situ", но и за счет аллохтонного сноса с более высоких участков. По периферии озерных котловин рост повторно-жильных льдов все время достаточно активен. Даже в мощных торфяниках крупные ледяные жилы нередки - мощные торфяники пронизанные ледяными жилами встречены автором в долинах р.р.Щучья, Юрибей, близ пос.Яптисале и в устье р.Сеяха (Зеленая). В тех частях торфяных массивов, где встречаются древесные горизонты крупные ледяные жилы, как правило, не обнаруживаются. Это вероятно, связано с локальными условиями увлажнения.
На спорово-пыльцевых диаграммах Ямала можно выделить последовательность максимумов пыльцы ивы затем - карликовой березки и чуть позже - ольховника, несмотря на то, что это время массового продвижения высокоствольной березы далеко на север.
3 этап: 7,8 - 4,6 тыс. лет назад. Примечательной ландшафтной особенностью этого периода является появление лиственницы в растительном покрове. Хотя древесина лиственницы относится к числу очень устойчивых к разложению пород [23], но лиственница на северном пределе своего существования имеет легко разлагающуюся древесину с рыхлой сердцевиной. Остатки северных лиственниц сохраняются только в исключительно благоприятных условиях. К тому же появление лиственницы в древостоях Ямала приходится на время частых пожаров. Поэтому отсутствие ее остатков в осадках нельзя интерпретировать как отсутствие в растительном покрове того времени.
Региональная составляющая на спорово-пыльцевых диаграммах, как правило, фиксирует максимальное содержание пыльцы ели, даже в самых северных районах Ямала появляется пыльца ели.
4 этап: 4,6 - 2,6 тыс. лет назад. Данный интервал характеризуется формированием ледяных жил в пойменных отложениях. Палиноспектры повторно-жильных льдов в разрезах высокой поймы р.Сеяха отражают региональный пыльцевой дождь в котором содержание пыльцы древесных снижается, в то время как концентрация и количество пыльцы типичных тундровых растений увеличивается. В составе палиноспектров с доминирует пыльца осок, злаков, а также карликовой березки и ольховника. В периоды усиления повторно-жильного образования на спорово-пыльцевых диаграммах высокой поймы отмечается повышение содержания пыльцы разнотравья. Вероятно с этим связан факт, отмеченный Р.М.Хантемировым и С.Г.Шиятовым [24] что значительное отступание границы хвойных пород к югу произошло около 3800 кал.лет назад, причем за сравнительно короткий промежуток времени. В течение последних 3800 кал. лет на Ямале наблюдалась самая низкая степень облесенности и наиболее южное положение северной границы редколесий. Однако, условия вегетационного периода были достаточно благоприятны, поэтому отмечается сравнительно высокая концентрация пыльцы тундровых растений –верескоцветных и осок
5 этап: 2,6 тыс. лет назад до современности. Практически во всех районах Ямала отмечаются условия, близкие к современным и палинологическая характеристика близкая к субфоссильным палиноспектрам. Выделение колебаний условий вегетационного периода возможно только на локальном уровне, как это сделано для территории Ямала в работе Ю.И. Прейс с соавторами [25]. Тем не менее отметим небольшое повышение содержания пыльцы сосны в разрезах низкой поймы и затем резкое снижение, что возможно отражает установленное отступание границы редколесий во временном интервале от 2350 до 2110 лет назад, зафиксированное дендрохронологическим методом [24].
Выводы:
· Разделение компонентов спектра на региональные и локальные составляющие, фиксирующие разномасштабные ландшафтные события оптимизирует анализ голоценовых спорово–пыльцевых диаграмм опорных разрезов повторно-жильных льдов Ямала. Региональные и дальнезаносные составляющие отражены также в палинологической характеристике повторно–жильных льдов.
· При анализе палиноспектров повторно-жильных льдов Ямала в качестве реперных в ряде случаев можно использовать палиноспектры отложений и льдов накопившихся в периоды похолоданий.
· Для интерпретации палинологических данных повторно-жильных льдов Ямала, важны не только температуры сезона вегетации, но и продолжительность сезона вегетации. Поэтому сумма положительных температур наиболее реальный показатель для реконструкций по палиноспектрам.
· Переотложение в группе древесных и кустарников существенно влияет на интерпретацию палиноспектров, поэтому адекватная оценка этого вида переотложения дает возможность сопоставления палиноспектров из разных разрезов. Это нивелирует различия палиноспектров, обусловленные разной степенью участия в них переотложенных палиноморф.
· В изученных голоценовых разрезах Ямала с синкриогенными повторно-жильными льдами зафиксированы среднепериодные контрастные изменения структуры спорово-пыльцевых спектров, выразившиеся в резкой смене палинодоминантов: палиноспектры с преобладанием пыльцы осок сменяются палиноспектры с преобладанием пыльцы злаков, с преобладанием полыни – доминированием маревых, с ольховником и карликовой березкой – палиноспектрами с пыльцой древовидной березы, с кедровый стланником – палиноспектрами с кедром сибирским, с большим содержанием недоразвитой пыльцы двудольных трав – спектрами с доминированием зеленых мхов. Основная периодичность смены палиноспектров происходит во временные интервалы длительностью 1,5-2 тыс. лет. Как правило, это связано с резким изменением условий вегетации, уровня базиса эрозии, повышением облачности в течение вегетационного периода, резким изменением направления и скорости ветра, влажности воздуха, различным временем формирования снежного покрова).
· Можно говорить об общих и локальных особенностях развития растительности, торфяников и повторно-жильных льдов в голоцене. Общие черты обусловлены глобальным изменением теплообеспеченности, в частности заметным увеличением температур вегетационного периода, при сохраняющихся суровых зимних условиях. Локальные черты связаны с неравномерным увлажнением территории, особенно важном факторе для криолитозоны, так как с этим связан и неравномерный характер промерзания массивов и различная интенсивность льдонакопления.
· Повторно-жильные льды формировались в течение всего голоцена на всей территории Ямала. В конкретных разрезах, можно выявить благоприятные условия для возникновения ледяных жил.
· Датирование отложений повторно-жильного комплекса связано с механизмом формирования отложений. При озерно-термокарстовом способе накопления осадков повторно-жильного комплекса пыльца и споры испытывают неоднократное переотложение. Нередко переотложение может даже превалировать над автохтонным накоплением палиноспектра. Определению масштаба и характера переотложения в этих случаях может содействовать палинологическая характеристика, с особым акцентом на выделение переотложенных палиноморф.
· Даже в верховых торфяниках и на задернованных поверхностях водоразделов Ямала содержание переотложенных палиноморф в среднем составляет 3-5%. Здесь пороговые значения содержания переотложенных палиноморф составляют около 15% от суммы подсчитанных пыльцевых зерен и спор.
· Палеоэкологическая информативность палиноспектров жильных льдов определяется возможностью реконструкций регионального пыльцевого дождя в датированный интервал времени.
· Спорово-пыльцевые спектры, заключенные в каком-либо прослое, можно рассматривать как суммарные для определенного временного интервала, предшествовавшего отложению этого прослоя. 14С датирование, очевидно, предпочтительно проводить по всем фракциям органики. В таком случае, наиболее молодая датировка даст наиболее точный возраст формирования отложений. Изменения спорово-пыльцевых спектров, отмечаемые в наблюдаемых льдистых разрезах, очевидно, отражают ход климатических колебаний в несколько сглаженном виде. В отдельных случаях в прибрежной зоне озер, где накапливается растительный детрит (который включен в разрез в виде прослоев аллохтонного торфа), обогащение осадков пыльцой и спорами может быть очень существенным по сравнению с синхронными палиноморфами. Приуроченности «богатых» спорово-пыльцевых комплексов к торфяным прослоям не наблюдается. При высокой концентрации аллохтонной органики анализ палиноспектров становится инструментом, который позволяет оценить датируемый материал.
Библиография
1. Голубева Л.В Спорово-пыльцевые спектры четвертичных отложений северо-западной части Западно-Сибирской низменности // Труды ГИН, вып. 31. 1960. С. 5–41.
2. Васильчук Ю.К., Петрова Е.А., Васильчук А.К. Некоторые черты палеогеографии голоцена Ямала // Бюллетень Комиссии по изучению четвертичного периода, вып. 52. 1983. С. 73–89.
3. Vasil'chuk Yu.K., Jungner H., Vasil'chuk A.C. 14C dating of peat and δ18O–δD in ground ice from Northwest Siberia // Radiocarbon. 2001. Vol. 43. N2B. P. 527–540.
4. Буданцева Н.А. Формирование повторно-жильных льдов и бугров пучения в голоцене (на примере Большеземельской тундры и севера Западной Сибири) / Диссертация на соискание учёной степени кандидата географических наук. М.: Моск ун-т. 2003. 205 с.
5. Васильчук Ю.К., Васильчук А.К. Мощные полигональные торфяники в зоне сплошного распространения многолетнемерзлых пород Западной Сибири (Thick polygonal peatlands in continuous permafrost of West Siberia) // Криосфера Земли. 2016. №4. C. 3–15.
6. Vasil'chuk Yu.K., van der Plicht J., Jungner H., Vasil'chuk A.C. AMS-dating of Late Pleistocene and Holocene syngenetic ice-wedges // Nuclear Instruments and Methods in Physics Research. Section B: Beam Interactions with Materials and Atoms. 2000. Vol.172. P. 637–641.
7. Кац В.Я. О динамике вечной мерзлоты в низовьях Оби в послеледниковое время // Бюллетень МОИП. Отдел биологич. 1939. Том 48. № 2-3. С. 44–56.
8. Тыртиков А.П. Динамика вечной мерзлоты вблизи ее южной границы в Западной Сибири в связи с развитием растительности // Природные условия Западной Сибири, вып.5. Под ред. А.И.Попова и В.Т.Трофимова.. М.: Изд-во Моск. ун-та. 1975. С. 99–122.
9. Тыртиков А.П. Лес на северном пределе в Азии. М. 1995. 144 с.
10. Вахта трехлистная. Menyanthes trifoliata L. // Атлас ареалов и ресурсов лекарственных растений СССР. М.: ГУГК. 1980. С. 16–17.
11. Кац В.Я., Федорова Р.В. О возможности использования спорово-пыльцевого анализа при проведении исследований в высокоширотных районах Арктики // Бюллетень МОИП. Отдел биологич. 1983. Том 88. №2. С. 63–70.
12. Попов А.И., Смирнова Т.И. Подземный лед в озерно-болотных отложениях на севере Западной Сибири // Вестник Моск. ун-та. Серия 5. География. 1964. №2. С. 32–39.
13. Данилов И.Д., Полякова Е.И. Криогенные и посткриогенные образования в аллювии и их палеогеографическое значение // Известия АН СССР. Серия географическая. 1978. № 2. С. 84–93.
14. Левковская Г.М. Зональные особенности современной растительности и рецентных спорово-пыльцевых спектров Западной Сибири // Методические вопросы палинологии: Труды III Междунар. палинол. конф. М., 1973. С. 116–120.
15. Васильчук А.К. Региональная и дальнезаносная пыльца в тундровых палиноспектрах // Известия Российской Академии Наук. Серия биологическая. 2005. №1. С. 85–99.
16. Фирсов Л.В., Троицкий С.Л., Левина Т.Н., Никитин В.П., Панычев В.А. Абсолютный возраст и первая для севера Западной Сибири стандартная пыльцевая диаграмма голоценового торфяника // Бюллетень комиссии по изучению четвертичного периода, вып. 41. М.: Наука. 1974. С. 121–127.
17. Васильчук Ю.К., Папеш В., Ранк Д., Сулержицкий Л.Д., Васильчук А.К., Буданцева Н.А., Чижова Ю.Н. Первые для севера Европы 14С-датированные изотопно-кислородная и дейтериевая диаграммы из повторно-жильного льда близ г.Воркуты // Доклады Российской Академии Наук. 2005. Том 400. №5. С. 684–689.
18. Арсланов Х.А., Каплянская Ф.А., Тарноградский В.Д., Тертычная Т.В. Радиоуглеродные датировки четвертичных отложений западного побережья п-ова Ямал // Бюллетень Комиссии по изучению четвертичного периода, вып. 55. 1986. С. 132–133.
19. Хотинский Н.А. Голоцен северной Евразии. Опыт трансконтинентальной корелляции этапов развития растительности и климата. М.: Наука. 1977. 198 с.
20. Ложкин А.В. Границы Берингии в позднем плейстоцене и голоцене // Четвертичная палеогеография Берингии. Магадан: СВКНИИ ДВО РАН. 2002. С. 4–12.
21. Украинцева В.В. Новые палинологические и палеоботанические свидетельства раннеголоценового потепления климата в высоких широтах Арктики // Ботанический журнал. 1990. Т. 75. № 1. С. 70–73.
22. Ritchie J.C., Cwynar L.C., Spear R.W. Evidence from northwest Canada for early Holocene Milancovitch maximum // Nature. 1983. Vol. 305. № 5930. P. 126–128.
23. Нащокин В.Д. Остатки ископаемой древесины из четвертичных отложений Сибири и значение их изучения для стратиграфии и палеогеографии // Четвертичный период Сибири. М.: Наука. 1966. С. 335–339.
24. Хантемиров Р.М., Шиятов С.Г. Радиоуглеродные и дендрохронологические датировки полуископаемой древесины на Ямале и их использование для изучения динамики лесотундровых экосистем // Биота Приуральской Субарктики в позднем плейстоцене и голоцене. Изд-во "Екатеринбург". 1999. С. 3–22.
25. Прейс Ю.И., Симонова Г.В., Слагода Е.А. Детальная стратиграфия и динамика хасырея Центрального Ямала в верхнем голоцене// Известия Томского политехнического университета. Инжиниринг георесурсов. 2016. Том 327. № 10. 35–49
References
1. Golubeva L.V Sporovo-pyl'tsevye spektry chetvertichnykh otlozhenii severo-zapadnoi chasti Zapadno-Sibirskoi nizmennosti // Trudy GIN, vyp. 31. 1960. S. 5–41.
2. Vasil'chuk Yu.K., Petrova E.A., Vasil'chuk A.K. Nekotorye cherty paleogeografii golotsena Yamala // Byulleten' Komissii po izucheniyu chetvertichnogo perioda, vyp. 52. 1983. S. 73–89.
3. Vasil'chuk Yu.K., Jungner H., Vasil'chuk A.C. 14C dating of peat and δ18O–δD in ground ice from Northwest Siberia // Radiocarbon. 2001. Vol. 43. N2B. P. 527–540.
4. Budantseva N.A. Formirovanie povtorno-zhil'nykh l'dov i bugrov pucheniya v golotsene (na primere Bol'shezemel'skoi tundry i severa Zapadnoi Sibiri) / Dissertatsiya na soiskanie uchenoi stepeni kandidata geograficheskikh nauk. M.: Mosk un-t. 2003. 205 s.
5. Vasil'chuk Yu.K., Vasil'chuk A.K. Moshchnye poligonal'nye torfyaniki v zone sploshnogo rasprostraneniya mnogoletnemerzlykh porod Zapadnoi Sibiri (Thick polygonal peatlands in continuous permafrost of West Siberia) // Kriosfera Zemli. 2016. №4. C. 3–15.
6. Vasil'chuk Yu.K., van der Plicht J., Jungner H., Vasil'chuk A.C. AMS-dating of Late Pleistocene and Holocene syngenetic ice-wedges // Nuclear Instruments and Methods in Physics Research. Section B: Beam Interactions with Materials and Atoms. 2000. Vol.172. P. 637–641.
7. Kats V.Ya. O dinamike vechnoi merzloty v nizov'yakh Obi v poslelednikovoe vremya // Byulleten' MOIP. Otdel biologich. 1939. Tom 48. № 2-3. S. 44–56.
8. Tyrtikov A.P. Dinamika vechnoi merzloty vblizi ee yuzhnoi granitsy v Zapadnoi Sibiri v svyazi s razvitiem rastitel'nosti // Prirodnye usloviya Zapadnoi Sibiri, vyp.5. Pod red. A.I.Popova i V.T.Trofimova.. M.: Izd-vo Mosk. un-ta. 1975. S. 99–122.
9. Tyrtikov A.P. Les na severnom predele v Azii. M. 1995. 144 s.
10. Vakhta trekhlistnaya. Menyanthes trifoliata L. // Atlas arealov i resursov lekarstvennykh rastenii SSSR. M.: GUGK. 1980. S. 16–17.
11. Kats V.Ya., Fedorova R.V. O vozmozhnosti ispol'zovaniya sporovo-pyl'tsevogo analiza pri provedenii issledovanii v vysokoshirotnykh raionakh Arktiki // Byulleten' MOIP. Otdel biologich. 1983. Tom 88. №2. S. 63–70.
12. Popov A.I., Smirnova T.I. Podzemnyi led v ozerno-bolotnykh otlozheniyakh na severe Zapadnoi Sibiri // Vestnik Mosk. un-ta. Seriya 5. Geografiya. 1964. №2. S. 32–39.
13. Danilov I.D., Polyakova E.I. Kriogennye i postkriogennye obrazovaniya v allyuvii i ikh paleogeograficheskoe znachenie // Izvestiya AN SSSR. Seriya geograficheskaya. 1978. № 2. S. 84–93.
14. Levkovskaya G.M. Zonal'nye osobennosti sovremennoi rastitel'nosti i retsentnykh sporovo-pyl'tsevykh spektrov Zapadnoi Sibiri // Metodicheskie voprosy palinologii: Trudy III Mezhdunar. palinol. konf. M., 1973. S. 116–120.
15. Vasil'chuk A.K. Regional'naya i dal'nezanosnaya pyl'tsa v tundrovykh palinospektrakh // Izvestiya Rossiiskoi Akademii Nauk. Seriya biologicheskaya. 2005. №1. S. 85–99.
16. Firsov L.V., Troitskii S.L., Levina T.N., Nikitin V.P., Panychev V.A. Absolyutnyi vozrast i pervaya dlya severa Zapadnoi Sibiri standartnaya pyl'tsevaya diagramma golotsenovogo torfyanika // Byulleten' komissii po izucheniyu chetvertichnogo perioda, vyp. 41. M.: Nauka. 1974. S. 121–127.
17. Vasil'chuk Yu.K., Papesh V., Rank D., Sulerzhitskii L.D., Vasil'chuk A.K., Budantseva N.A., Chizhova Yu.N. Pervye dlya severa Evropy 14S-datirovannye izotopno-kislorodnaya i deiterievaya diagrammy iz povtorno-zhil'nogo l'da bliz g.Vorkuty // Doklady Rossiiskoi Akademii Nauk. 2005. Tom 400. №5. S. 684–689.
18. Arslanov Kh.A., Kaplyanskaya F.A., Tarnogradskii V.D., Tertychnaya T.V. Radiouglerodnye datirovki chetvertichnykh otlozhenii zapadnogo poberezh'ya p-ova Yamal // Byulleten' Komissii po izucheniyu chetvertichnogo perioda, vyp. 55. 1986. S. 132–133.
19. Khotinskii N.A. Golotsen severnoi Evrazii. Opyt transkontinental'noi korellyatsii etapov razvitiya rastitel'nosti i klimata. M.: Nauka. 1977. 198 s.
20. Lozhkin A.V. Granitsy Beringii v pozdnem pleistotsene i golotsene // Chetvertichnaya paleogeografiya Beringii. Magadan: SVKNII DVO RAN. 2002. S. 4–12.
21. Ukraintseva V.V. Novye palinologicheskie i paleobotanicheskie svidetel'stva rannegolotsenovogo potepleniya klimata v vysokikh shirotakh Arktiki // Botanicheskii zhurnal. 1990. T. 75. № 1. S. 70–73.
22. Ritchie J.C., Cwynar L.C., Spear R.W. Evidence from northwest Canada for early Holocene Milancovitch maximum // Nature. 1983. Vol. 305. № 5930. P. 126–128.
23. Nashchokin V.D. Ostatki iskopaemoi drevesiny iz chetvertichnykh otlozhenii Sibiri i znachenie ikh izucheniya dlya stratigrafii i paleogeografii // Chetvertichnyi period Sibiri. M.: Nauka. 1966. S. 335–339.
24. Khantemirov R.M., Shiyatov S.G. Radiouglerodnye i dendrokhronologicheskie datirovki poluiskopaemoi drevesiny na Yamale i ikh ispol'zovanie dlya izucheniya dinamiki lesotundrovykh ekosistem // Biota Priural'skoi Subarktiki v pozdnem pleistotsene i golotsene. Izd-vo "Ekaterinburg". 1999. S. 3–22.
25. Preis Yu.I., Simonova G.V., Slagoda E.A. Detal'naya stratigrafiya i dinamika khasyreya Tsentral'nogo Yamala v verkhnem golotsene// Izvestiya Tomskogo politekhnicheskogo universiteta. Inzhiniring georesursov. 2016. Tom 327. № 10. 35–49
|
 Рус
Рус











